
Abstract
Phased telomere-to-telomere (T2T) genome assemblies are revolutionizing our understanding of long-hidden genome biology “dark matter” such as centromeres, rDNA repeats, inter-haplotype variation, and allele-specific expression (ASE), yet insights into dikaryotic fungi that separate their haploid genomes into distinct nuclei are limited. Here, we explore the impact of dikaryotism on the genome biology of a long-term asexual clone of the wheat pathogenic fungus Puccinia striiformis f. sp. tritici. We use Oxford Nanopore Technologies (ONT) duplex sequencing combined with Hi-C to generate a T2T nuclear-phased assembly with >99.999% consensus accuracy. We show that this fungus has large regional centromeres enriched in LTR retrotransposons, with a single centromeric dip in methylation that suggests one kinetochore attachment site per chromosome. The centromeres of homologous chromosomes are most often highly diverse in sequence, and kinetochore attachment sites are not always positionally conserved. Each nucleus carries a unique array of rDNAs with more than 200 copies that harbor nucleus-specific sequence variations. The inter-haplotype diversity between the two nuclear genomes is shaped by large-scale structural variations linked to transposable elements. ONT long-read cDNA analysis across dormancy and distinct host infection conditions revealed pervasive ASE for ∼20% of the heterozygous genes. Genes encoding secreted proteins, including putative virulence effectors, are significantly enriched in ASE genes that appear to be linked to elevated CpG gene body methylation of the lower-expressed allele. This suggests that epigenetically regulated ASE is likely a previously overlooked mechanism facilitating plant infection. Overall, our study reveals how dikaryotism uniquely shapes key eukaryotic genome features.
Telomere-to-telomere (T2T) and haplotype-resolved genome assemblies have become the norm in eukaryotic genomics with advances in long-read sequencing technologies. Complete genome assemblies are fundamental for addressing key questions in genome biology that were previously hidden in the “dark matter” of genomes. Key breakthroughs have revolved around centromeres and the embedded kinetochore attachment sites pivotal for understanding karyotype diversity and evolution (Logsdon et al. 2024; Mastrorosa et al. 2024), as well as the notoriously repetitive ribosomal DNA (rDNA) arrays whose sequences have only been recently completed in humans (Nurk et al. 2022) and Arabidopsis (Fultz et al. 2023). Full haplotype resolution enables precise characterization of inter-haplotype variations predominantly shaped by heterozygosity, structural rearrangements, and transposable element (TE) movements (Gluck-Thaler et al. 2022; Hartmann 2022; Ferguson et al. 2024). Further, the complete picture of allelic information facilitates robust assessment of allele-specific expression (ASE) uncovering the underlying variations in cis-regulatory elements and epigenetic regulation, which can have important implications on phenotypic variability, as well established in mammals and plants (e.g., Shao et al. 2019; Cleary and Seoighe 2021; St. Pierre et al. 2022; Tian et al. 2022; Shi et al. 2024).
Although there have been important novel insights into diploid and polyploid genome organization, especially in plants (e.g., Belser et al. 2021; Hu et al. 2021; Sun et al. 2022), less is known about the fungi-specific dikaryotic state in which two haploid genomes are contained in separate nuclei in the same cytoplasm and propagated in a coordinated manner during cell division (Anderson and Kohn 2014; Li et al. 2019; Sperschneider et al. 2023a,b). Dikaryotism is highly successful, with an estimated 400,000 taxa in the fungal subphylum Basidiomycota relying on it for significant time periods, for example, during fruiting body formation in mushrooms and infection processes in rusts and smuts (Schmidt-Dannert 2016; James et al. 2020). Early partially phased assemblies of rust fungi genomes indicated high levels of heterozygosity and the presence/absence polymorphisms between the nuclear genomes; however, they lacked the resolution to identify individual haplotypes (Cuomo et al. 2017; Schwessinger et al. 2018, 2020; Vasquez-Gross et al. 2020). The latest fully haplotype-phased and nuclear-assigned T2T genomes have provided the first insights into organization of genes involved in mating behavior and contributed to our understanding of reproductive mechanisms generating novel genetic diversity (Schwessinger et al. 2022; Li et al. 2023a; Luo et al. 2024). This includes the somatic exchanges of nuclei between asexual rust lineages that are adapted to the infection of cereals (Sperschneider et al. 2023a; Henningsen et al. 2024), and sexual recombination in permissive hosts (Rodriguez-Algaba et al. 2022; Wang et al. 2022; Du et al. 2023). Such genetic reshuffling produces novel allele combinations of so-called avirulence (Avr) genes that encode secreted effector proteins essential for host infection. Rust effectors are under strong selection pressure to diversify into nonrecognized virulence alleles because they can be recognized by cognate immune receptors encoded by wheat resistance genes (Chen et al. 2017; Salcedo et al. 2017; Ortiz et al. 2022). This is necessary to escape the plant immune system and to confer the ability to infect new host varieties.
Recent progress in heterokaryotic fungi genomics, such as in arbuscular mycorrhizal fungi, has revealed extensive nuclear genome variations in structural and gene content (Li et al. 2019; Sperschneider et al. 2023b), as well as nuclear-level transcriptomic and epigenetic differences similar to findings in button mushroom Agaricus bisporus (Gehrmann et al. 2018), although key questions remain around the impact of dikaryotic genome organization on the evolution of centromeres, rDNA repeats, detailed inter-haplotype variations, ASE and methylation differences at locus resolution. Here, we use a long-term asexual clone of the wheat stripe rust fungus Puccinia striiformis f. sp. tritici (Pst) dating back at least 80 years to address these questions in a globally important wheat pathogen (Wellings 2007; Thach et al. 2016; Schwessinger et al. 2018). We leveraged high-accuracy Oxford Nanopore Technologies (ONT) duplex long reads to generate the first ONT-based T2T, fully nuclear-phased genome assembly for a dikaryotic fungus. We combined this with comprehensive ONT long-read cDNA data sets sampled during dormancy and host pathogenesis for high-quality gene annotations and differential expression analysis at gene- and allele-specific levels. Our study sheds new light onto the genome biology and adaptive evolutionary potential of rust fungi, with important implications for managing their agricultural impacts.
Results
A T2T nuclear-phased genome assembly of Pst104E based on high-quality ONT long-read sequencing
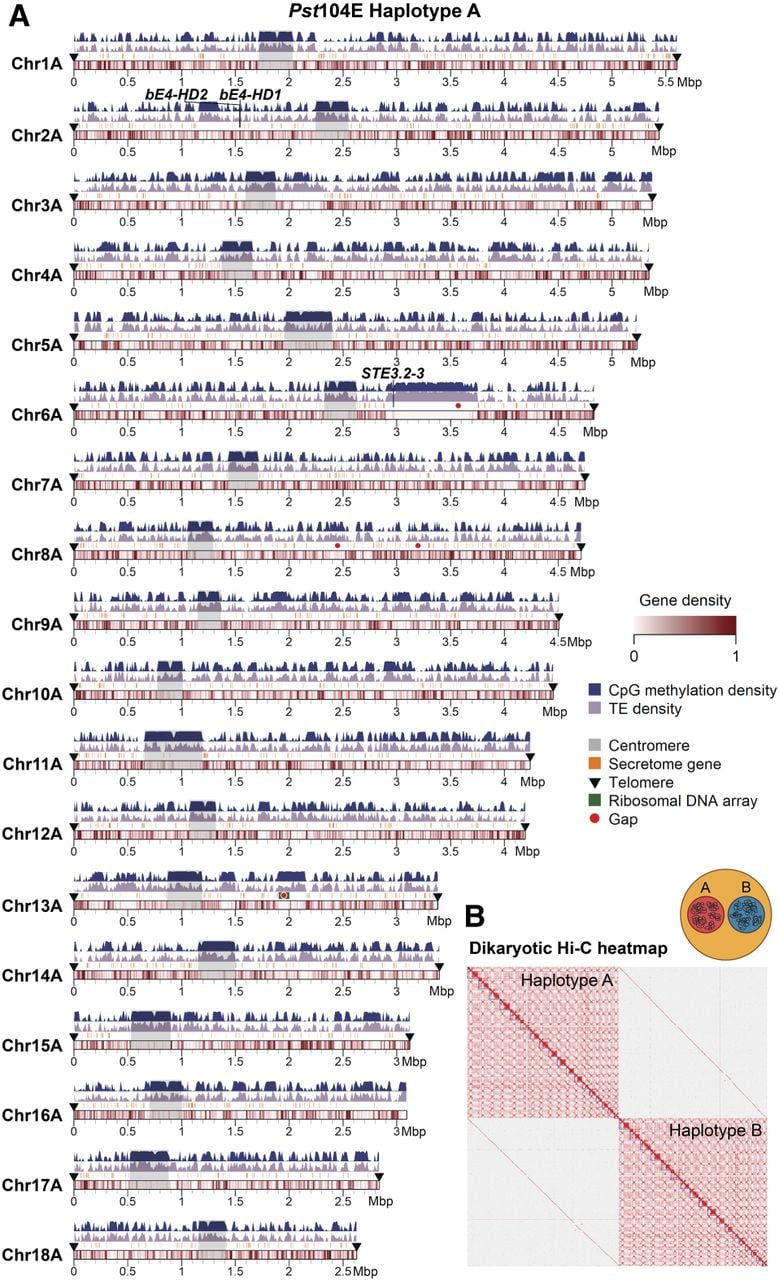
We generated a dikaryotic Pst genome assembly for a representative isolate of the Australian founder pathotype 104E137A− (abbreviated as Pst104E), which belongs to the long-term asexual PstS0 lineage (Wellings 2007; Schwessinger et al. 2018, 2020). We assembled ONT duplex reads combined with ultralong simplex reads and Hi-C (Supplemental Table S1), followed by scaffolding and manual curation. This resulted in 36 chromosome assemblies corresponding to the 18 homologous pairs, which we sorted by average length of both haplotypes (Chr 1 to Chr 18) (Fig. 1A; Table 1; Supplemental Fig. S1). Of these, 35 were assembled T2T, and each telomere had about 43 repeats on average, consistent with other basidiomycetes (Ramírez Nasto et al. 2011; Schwessinger et al. 2020; Sperschneider et al. 2021). The Hi-C contact heatmap clearly grouped the homologous chromosomes into two nuclear complements as expected for dikaryons (Fig. 1B). Five remaining gaps were found at repetitive regions, for example, a ∼650 kbp TE-rich region on Chr 6A near the previously identified mating type PR locus (Supplemental Fig. S2; Luo et al. 2024), as well as two rDNA arrays on Chr 13A and Chr 13B (Supplemental Fig. S3).
A nuclear-phased, chromosome-scale genome assembly of the dikaryotic fungus Puccinia striiformis f. sp. tritici (Pst) isolate 104E. (A) Karyoplot of the 18 chromosomes of haplotype A, showing CpG methylation and transposable element (TE) density as peaks, and gene density as heatmaps within chromosome ideograms (10 kbp sliding windows). Locations of centromeres, telomeres, secretome genes, the ribosomal DNA (rDNA) array, and assembly gaps are annotated as per the legend. The mating type loci are labeled in black text above the corresponding chromosomes. (B) Hi-C contact heatmap of the full dikaryotic genome assembly consisting of 36 chromosomes. The two nuclear haplotypes A and B display a clear signal of spatial separation.
Summary of assembly statistics and quality metrics of the ONT duplex genome assembly of Pst104E
| Statistic | Full dikaryotic | Haplotype A | Haplotype B |
|---|---|---|---|
| Assembly size (bp) | 152,328,638 | 77,128,783 | 75,199,855 |
| No. of T2T chromosomes | 35/36 | 17/18 | 18/18 |
| N50 (bp) | 4,614,849 | 4,713,886 | 4,606,998 |
| % GC content | 44.41 | 44.41 | 44.42 |
| % TE content | 44.51 (68 Mbp) | 45.16 (35 Mbp) | 43.87 (33 Mbp) |
| No. of gaps | 5 | 4 | 1 |
| % complete BUSCOs | 92.6 | 91.9 | 91.4 |
| No. of heterozygous SNPs/Mbp | 6.8 | 6.9 | 6.7 |
| No. of homozygous SNPs/Mbp | 1.1 | 1.3 | 0.8 |
| LAI | 18.3 | 27.2 | 24.8 |
| CRAQ R-AQI | 94.7 | 94.3 | 95.1 |
| CRAQ S-AQI | 97.4 | 97.4 | 97.4 |
We evaluated the quality of the final curated assembly. The full assembly yielded 92.6% of complete BUSCOs (Manni et al. 2021). We used the long terminal repeat (LTR) assembly index (LAI) (Ou et al. 2018) to assess contiguity at repetitive LTR retrotransposons. Haplotype A and B assemblies each received LAI score of 27.2 and 24.8, classifying them to the highest rank based on well-assembled repeats. The per-base accuracy Phred score was 57.4 corresponding to >99.999% consensus accuracy, in line with recent Pacific Biosciences (PacBio) HiFi assemblies (Li et al. 2023a; Sperschneider et al. 2023a; Wang et al. 2024). CRAQ (Li et al. 2023b) was used to detect clipped alignments indicative of regional and structural errors that could be computed into R/S–assembly quality indices (AQIs). Our assembly achieved R/S-AQIs of 94.7 and 97.4, both meeting reference quality. Rare residual errors were reflected by the low density of SNPs detected as homozygous (1.1/Mbp, 161 total) and heterozygous (6.8/Mbp, 1038 total).
Analysis of the mapping read depth supported full haplotype phasing with a single peak corresponding to the expected haploid 1× depth (Supplemental Fig. S4). To detect putative phase switch errors, we quantified Hi-C paired alignments within and between haplotypes. About 99.2% of the Hi-C mappings were between chromosomes contained within one nucleus (within-haplotype links), with 0.8% linking chromosomes contained in separate nuclei (cross-haplotype links) (Table 2; Supplemental Fig. S5). These results demonstrate a highly complete and accurate nuclear-phased genome assembly of Pst104E.
Hi-C contact statistics confirming phasing correctness of the Pst104E genome assembly
| Types of Hi-C contacts (MAPQ ≥ 20) | Counts |
|---|---|
| Total Hi-C links | 1,126,566 |
| Within-haplotype links | 1,117,107 (99.2%) |
| cis-chromosome | |
| Haplotype A | 479,583 (42.6%) |
| Haplotype B | 467,937 (41.5%) |
| trans-chromosome | |
| Haplotype A | 85,667 (7.6%) |
| Haplotype B | 83,920 (7.5%) |
| Cross-haplotype links | 9459 (0.8%) |
ONT cDNA sequencing enables high-quality evidence-guided gene annotations
We aimed to improve on current fungal gene annotations by incorporating extensive long-read cDNA sequencing data sets (Pardo-Palacios et al. 2024). We generated a detailed time course of ONT direct cDNA data sets for Pst104E gene annotation. The transcripts were sampled from six conditions with four replicates each (Supplemental Table S2). These included ungerminated (UG) urediniospores as the dormancy control and infected wheat leaf tissues at 4, 6, 8, 10, and 12 days post infection (dpi). Principal component analysis (PCA) demonstrated clear clustering of the technical replicates of each sample, with separation of samples representing dormancy (UG) and macroscopically asymptomatic (4, 6, and 8 dpi) and symptomatic (10 and 12 dpi) stages (Supplemental Fig. S6). We also complemented gene annotation with multiple publicly available Illumina RNA-seq data sets (Supplemental Table S3; Dobon et al. 2016; Schwessinger et al. 2018; Zhao et al. 2021). In total, we annotated 15,142 protein-coding genes on haplotype A and 14,938 on haplotype B, improving the complete BUSCO score to 94.8%. Functional annotation identified ∼15% of the gene models that encode secreted proteins without predicted transmembrane domain.
ONT direct cDNA sequencing has demonstrated quantitative power consistent with short-read RNA-seq, making it suitable for differential expression analysis (Sessegolo et al. 2019; Grünberger et al. 2022; Pardo-Palacios et al. 2024). We performed differential expression analysis with our ONT cDNA data sets to identify candidate Avr effector genes. We searched for secretome genes that are upregulated early in wheat infection (4, 6, and 8 dpi) relative to UG, as their functions likely correlate with pathogenesis. A total of 1318 secretome genes were found to be upregulated early during infection. These were shortlisted to hemizygous genes (single copy) for reduced functional redundancy, resulting in 97 high-priority candidates for future functional validation (Supplemental Table S4).
TE annotations
TEs are major components of genomes of many basidiomycetes (Castanera et al. 2017; Corre et al. 2025). TE annotations for our Pst104E assembly revealed that both haploid genomes shared similar TE content and composition, covering 44.51% of the genome space (Table 1). This was consistent with findings for other published Pst genomes (Zheng et al. 2013; Schwessinger et al. 2018, 2022). Class I (retrotransposons) and Class II (DNA transposons) accounted for 15% and 18.7%, respectively, of the genome (Supplemental Table S5). LTRs comprised the most abundant retrotransposons, predominated by the Ty3/Gypsy. Terminal inverted repeats (TIRs) represented the majority of DNA transposons.
Pst centromeres are highly diverse and enriched with LTR retrotransposons
We set out to identify Pst104E's centromeric regions and analyzed their sequence composition. Basidiomycete centromeres are often characterized by hypermethylated TE-rich and gene-poor regions (Guin et al. 2020). We estimated their positions from the Hi-C heatmap based on the strong inter-chromosomal contact peaks (“bowtie” shapes) caused by Rabl centromere clustering (Fig. 1B; Varoquaux et al. 2015; Muller et al. 2019). These interaction sites were enriched for TEs and overlapped with gene-sparse regions spanning hundreds of kilobases (Figs. 1A, 2A). Using ONT-derived methylation data, we found substantially more methylated CpGs within all 36 inferred centromeric regions, Cen1A to Cen18B (94.9%–98.3%), than in noncentromeric regions (32.9%–45.3%) (Fig. 2B).
Pst104E centromeres are highly diverse with haplotype-specific sequences and are enriched in retrotransposons. (A) Percentage of TE coverage in centromere and noncentromere regions of Pst104E. Each dot represents one of the 36 chromosomes. Student's t-test: (***) P < 0.001. (B) Percentage of coverage of methylated CpG sites in centromere and noncentromere regions of Pst104E. Each dot represents one of the 36 Pst104E chromosomes. Student's t-test: (***) P < 0.001. (C) Size ratio of haplotype A centromeres compared to haplotype B centromeres. Outlier ratios exceeding 1.5 times the interquartile range are highlighted in red. (D) Pairwise alignment dotplot between centromeres of haplotypes A and B. Only alignment blocks >100 bp with a minimum of 90% sequence identity are shown. Each color denotes an alignment type: blue, unique forward alignments; green, unique reverse alignments; and orange, repetitive alignments. (E) Enrichment of TE superfamilies within centromeres compared with noncentromere regions. Statistical significance was assessed using permutation tests on each chromosome. Only abundant TE superfamilies with >1% of total genome coverage are shown. The color scalebar denotes the test statistic, defined as the observed difference in the percentage of TE coverage within and outside centromeres. P-values represent the proportion of permuted values equal to or more extreme than observed. FDR < 5% was applied to correct for multiple testing: (*) P < 0.05, (**) P < 0.01, (****) P < 0.0001.
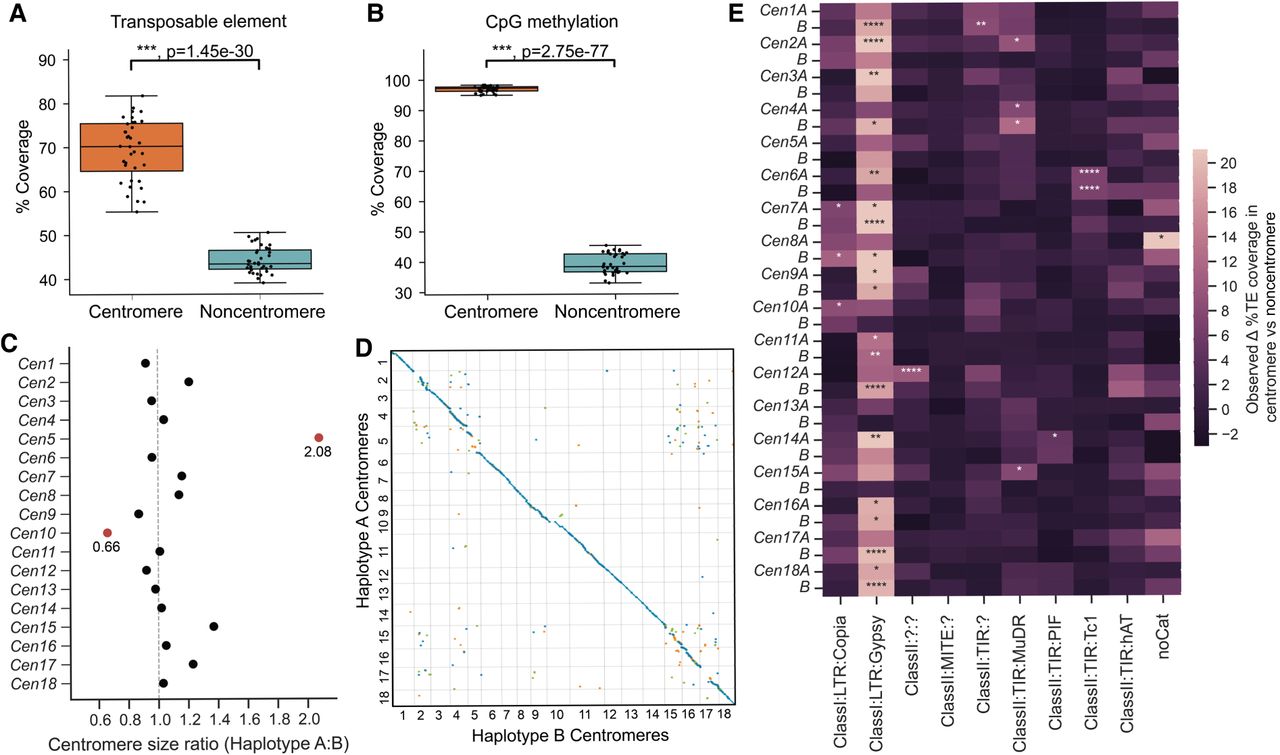
Pst104E centromere sizes varied approximately 2.5-fold between 210 and 538 kbp (mean 304 kbp) (Supplemental Table S6), categorizing them as “large regional” centromeres that are known to support multiple spindle microtubule attachments (Bodor et al. 2014; Yadav et al. 2018a). Most homologous chromosomes shared similar centromere lengths with differences under 1.4-fold, whereas Cen5A stood out being double the length of Cen5B (Fig. 2C). Pairwise alignments of centromeric regions revealed varying levels of macrocollinearity between haplotypes, ranging from almost complete (e.g., Cen3, -12, -13, and -18) to negligible synteny (e.g., Cen2, -5, -15, and -17) (Fig. 2D). Given the high TE density, such a diverse range in centromeric sequence conservation prompted us to examine their TE composition in search of elements possibly linked to centromere function.
We conducted permutation tests to determine the enrichment of abundant TE superfamilies within each Pst centromere compared with noncentromeric regions (Fig. 2E; Supplemental Table S7). Most centromeres (25 out of 36) were found to be significantly enriched for one to two TE superfamilies that belonged to retro- and/or DNA transposons (Fig. 2E). Of these, 19 centromeres were significantly enriched for Ty3/Gypsy LTRs, occasionally cocolonized by TIRs. Given the balanced representation of retro- and DNA transposons throughout the genome, this finding suggests that retrotransposons might have a more prominent role in Pst centromere formation than DNA transposons.
Centromeres contain a single putative kinetochore attachment site
It is unclear if rust fungi have one or multiple kinetochore attachment sites given they have “large regional” centromeres (Fig. 2; Yadav et al. 2018a; Sperschneider et al. 2021). Kinetochores typically assemble at a hypomethylated stretch of DNA embedded within the centromere termed the “centromere dip region” (CDR), which is marked by the centromere-specific histone variant CENP-A (Akiyoshi 2019; Sundararajan and Straight 2022; Logsdon et al. 2024). We examined the CpG methylation pattern along the Pst104E centromeres to locate CDRs as a qualitative proxy for potential kinetochore sites. Throughout all centromeres, we consistently observed a single methylation depletion “valley” spanning 24.8 kbp on average, which is characteristic of CDR (Fig. 3A; Supplemental Fig. S7; Supplemental Table S6). The CDR lengths were similar between haplotypes (Fig. 3B). The positioning of CDRs were generally conserved (Figs. 3C, 2D). The two notable exceptions were Cen3 and Cen13, whose CDR haplotypes were placed further than 20% of their centromere lengths apart (Fig. 3C).
Analysis of centromere dip regions (CDRs) within Pst104E centromeres suggest a single putative kinetochore attachment site per chromosome. (A) CpG methylation profiles (black histograms) and percentage of AT content (red lines) across Pst104E centromeres Cen1A to Cen9A as examples. A consistent methylation depletion valley with a mean size of ∼24.8 kbp was observed throughout all centromeres, indicating CDR signals (marked by gray dotted line). (B) Estimated sizes of the CDRs for each chromosome in haplotype A and B. Student's t-test: (NS) P > 0.05. (C) Differences in the relative positions of CDRs between haplotypes for each homologous chromosome pair. Differences that exceed 0.2 are highlighted in red. (D) Sequence alignment dotplot between Cen3A and Cen3B and their respective CpG methylation profiles (black histograms). The dotted red box highlights sequence divergence between Cen3A and Cen3B corresponding to the Cen3B CDR. (E) Detailed synteny analysis of the region highlighted in D. Long orange arrows indicate copies of hapB-B-G1437-Map9 belonging to the Ty3/Gypsy LTR retrotransposon superfamily. Gray shading indicates homologous sequences with percentage identity shown in boxes. Numbers above and below the orange arrows indicate the respective percentage of methylated CpGs.
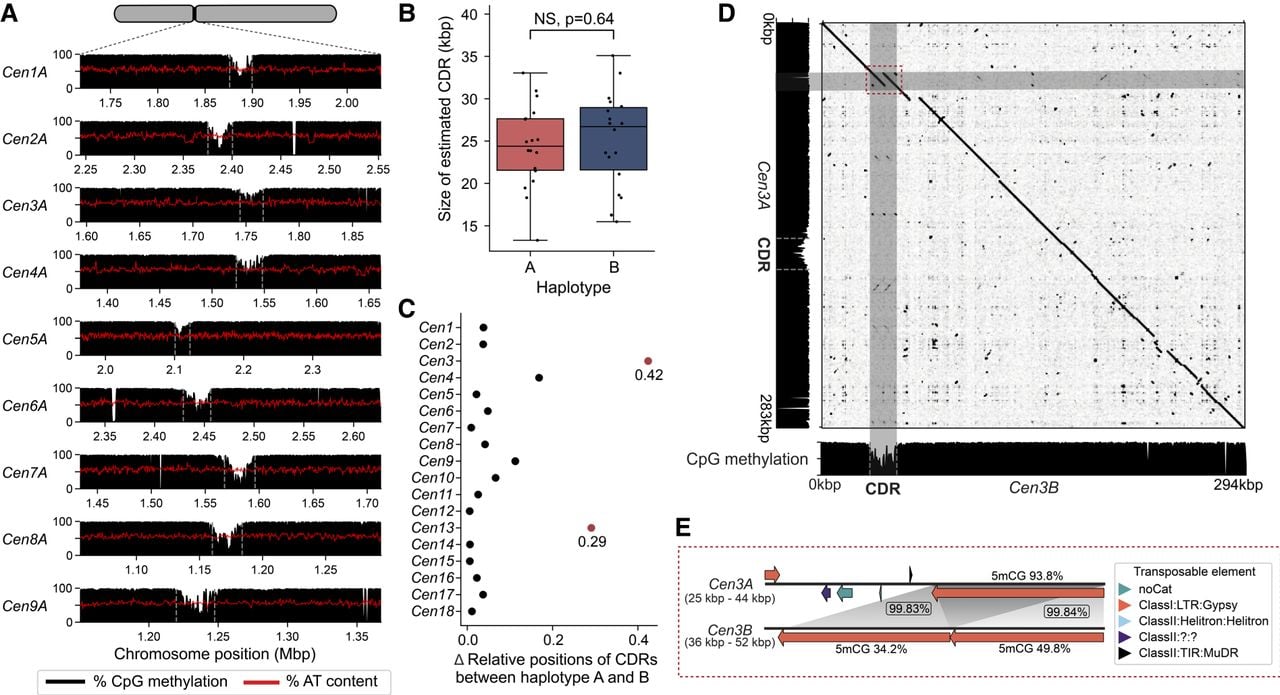
We next investigated whether the CDR signals were linked to increased AT content as reported for other fungi (Yadav et al. 2018a; Sankaranarayanan et al. 2020; Narayanan et al. 2024). No differences in AT content were detected when comparing CDRs with centromeric or noncentromeric regions (Supplemental Fig. S8). The only exception was Cen3B CDR, which has an elevated AT content of 59.8% compared with the rest of the centromere and the overall genome average of 55.6%. Close investigation revealed two nearly identical AT-rich copies of a Ty3/Gypsy retrotransposon family (hapB-B-G1437-Map9), together covering 91% of Cen3B CDR. Sequence alignment revealed that the corresponding syntenic region on Cen3A contained only one copy of this TE family, which was not involved in CDR formation (Fig. 3D). Consistently, this single copy on Cen3A was highly methylated on its CpG sites (93.8%), whereas the two TE copies on Cen3B CDR were lowly methylated (34.7% and 49.8%) (Fig. 3E). This was despite >99.5% identity shared by the three TE copies. These results highlight that centromere and kinetochore attachment site formation is not solely driven by primary DNA sequence composition.
Nucleus-specific variations in the rDNA arrays
We investigated the rDNA composition in Pst104E to better understand its dynamics in the context of a dikaryotic genome with physical separation of the haploid genomes. Pst104E has a single rDNA cluster per nuclear genome on the q-arm of Chr 13 (Fig. 1A). Both haplotypes were incompletely assembled with a gap at the rDNA cluster, indicating an underrepresentation of this complex locus (Supplemental Fig. S3). The assembled rDNA copies were oriented with transcription directed away from the centromere and toward the telomere (Fig. 4A). We reconstructed the canonical rDNA repeat (see Methods), which included the 45S transcription unit (18S, 5.8S and 25S rRNA genes) separated by two internal transcribed spacers (ITS1 and ITS2), followed by two intergenic spacers (IGS1 and IGS2) and a 5S rRNA gene that is barely transcribed in between (Fig. 4B; Supplemental Fig. S9). We defined 18S and IGS2 as the start and end of an rDNA unit for the subsequent analysis.
The two dikaryotic nuclear haplotypes of Pst104E harbor distinct rDNA subtypes. (A) Schematic view of the rDNA tandem repeat array located on Pst104E Chr 13. (B) Diagram of a single canonical rDNA unit of Pst104E containing the transcription start site in the 5′ external transcribed spacer, the catalytic rRNA genes (18S, 5.8S, 25S, and 5S), two internal transcribed spacers (ITS1 and ITS2), and two intergenic spacers (IGS1 and IGS2). The alignment shows two dominant rDNA subtypes (1 and 2), which differ by SNPs and structural variations in both IGS. (C) Estimated copy number of rDNA subtypes 1 and 2 based on ONT duplex and Illumina read data sets. (D) Workflow of our rDNA Hi-C analysis designed to determine the physical location of the two dominant rDNA subtypes in each nuclear haplotype. (E) Mapping quality (MAPQ) distributions of Hi-C reads whose paired mates contained rDNA subtype-specific or non-subtype-specific 31-mers, categorized by the nuclear haplotype they mapped against.
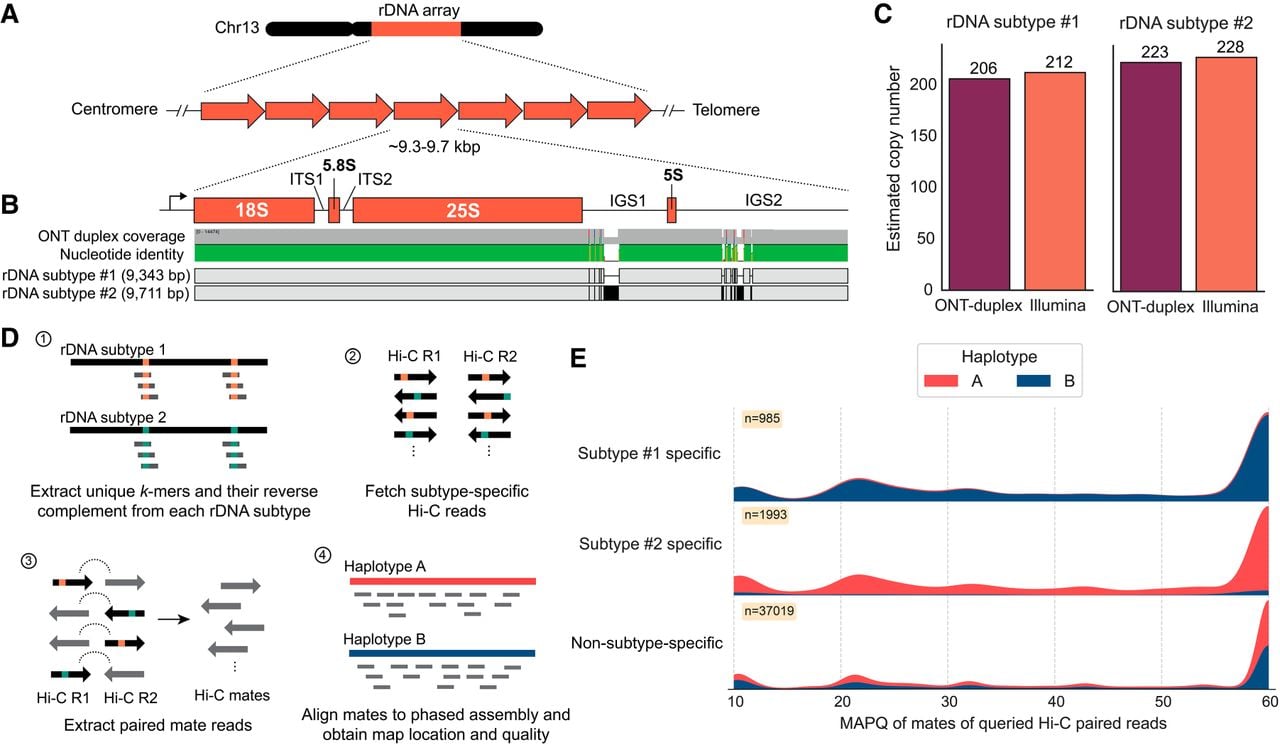
Detailed analyses of sequencing reads revealed two major rDNA subtypes (1 and 2) supported by substantial read coverage at near-equal frequencies. The estimated copy number ranged from 206 to 212 copies for rDNA subtype 1 and 223 to 228 copies for subtype 2, totaling about 434 copies (Fig. 4C). The two subtypes were 9343 bp and 9711 bp in length, with polymorphisms contained in the repeats nested within IGS1 and IGS2 (96.1% sequence identity) (Supplemental Fig. S10). We also identified 12 variants of the two subtypes (1.1–1.9 and 2.1–2.3) based on low-frequency SNP analysis (Supplemental Fig. S11; Supplemental Table S8). Most of these SNPs occurred in the 18S gene. The subtype variants 1.1–1.9 and 2.1–2.3 collectively accounted for ∼23% of the total copy number, suggesting incomplete rDNA homogenization (Supplemental Table S8).
Given the long clonal history of Pst104E (Schwessinger et al. 2020), the individual haplotypes are expected to have been stably inherited in separate nuclei without karyogamy or meiotic crossovers. We therefore hypothesized that the sequence variations in the two dominant rDNA subtypes might be nucleus specific. To test this, we took advantage of Hi-C read pairs that contained rDNA subtype-specific k-mers and asked where the alternate read mate pair mapped (Fig. 4D). About 94.3% of mates associated with rDNA subtype 1 mapped to haplotype B, whereas 90.2% of those associated with subtype 2 mapped to haplotype A (Fig. 4E); the remaining cross-haplotype links were likely technical noises owing to some less informative subtype-specific k-mers (such as those derived from the IGS2 minisatellite-like repeats) and sequencing artifacts. The control procedure, which involved aligning subtype-nonspecific rDNA Hi-C read pairs, showed a near-equal proportion of Hi-C mates mapped against each haplotype. Together, this implies that each rDNA subtype was associated with a different haplotype and that each nuclear genome contains its own major rDNA subtype array.
Large-scale structural variations driven by TEs shape inter-haplotype diversity
The phased Pst104E assembly allowed us to assess its inter-haplotype structural variations (SVs). The haplotypes were 78.8% syntenic (Fig. 5A), with 60.2 Mbp of haplotype A (78.0%) and 60.1 Mbp of haplotype B (79.6%) identified as highly continuous syntenic blocks (Fig. 5B). All identified SV types (duplications, large indels, translocations, and inversions) collectively occupied ∼11% and 9%, respectively, of the total lengths of haplotypes A and B (Fig. 5B; Supplemental Table S9). About 10% of each haplotype's sequence was not alignable and was therefore hemizygous. Syntenic regions had the largest average length, with most ranging 10–100 kb. The different SV types had various length distributions mostly centered at ∼1–10 kb (Fig. 5B). We used permutations to test if the SVs and their 2 kbp flanking regions were enriched for specific genomic features. This revealed a significant enrichment of TEs across all analyzed SV types, especially duplications, possibly owing to the replicative mechanism of retrotransposons (Fig. 5C; Supplemental Table S10). In contrast, protein-coding genes were depleted at SVs.
Pst104E’s inter-haplotype variation is shaped by large-scale structural variation (SV) and TEs. (A) Synteny and structural rearrangements between chromosome pairs of the two nuclear haplotypes, with A as reference and B as query. (B) Length distribution (left) of different SV types, including syntenic and unaligned regions. Counts of each SV type are shown. The bar chart (right) represents the total genome length covered by each sequence category. (C) Enrichment (red) and depletion (blue) of different genomic features (TEs and genes) within SVs including ±2 kbp flanking regions tested using permutations. The color scale denotes the observed difference in the percentage of coverage of a genomic feature located within and outside of a given set of SVs. P-values indicate the proportion of permuted coverage values less than, equal to, or greater than the observed: (*) P < 0.05, (**) P<0.01, (***) P<0.001, FDR-corrected. (D) Flowchart summarizing the classification of hemizygous, heterozygous, and homozygous protein-coding genes and their further categorization. Hemizygous genes were intersected with secretome genes upregulated during infection time points (4, 6, or 8 dpi) to shortlist Avr candidates. Heterozygous biallelic genes were used for allele-specific expression (ASE) analysis.
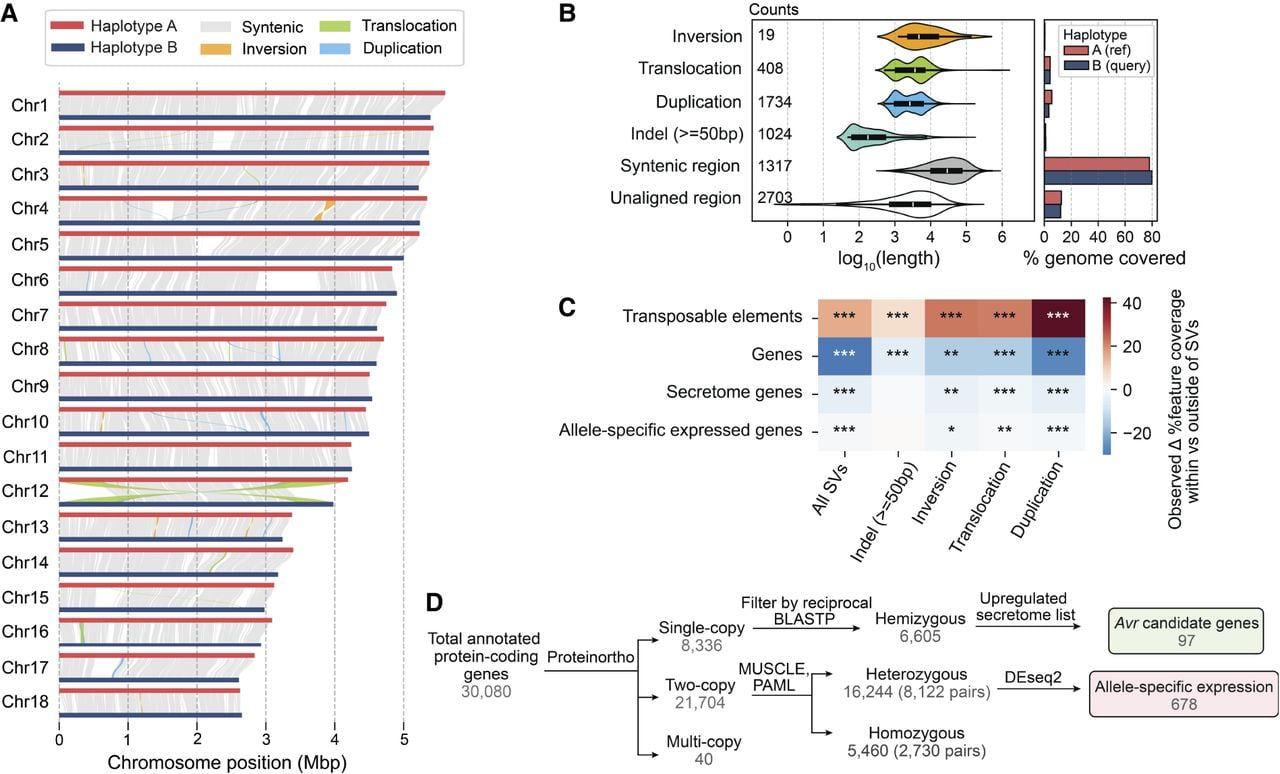
Next, we assessed the impact of inter-haplotype variation on genes by analyzing their sequence conservation and synteny (Fig. 5D). We robustly identified 3391 and 3214 hemizygous genes unique to either haplotype A or B, respectively. A total of 21,744 genes shared at least one homolog in the alternative haplotype. We analyzed one-to-one gene pairs and performed codon-aware alignment to compute their divergence values at synonymous and nonsynonymous sites (Supplemental Table S11), identifying 2730 homozygous pairs. The remaining 8122 displayed divergence greater than zero and were therefore defined as heterozygous biallelic pairs.
ASE is correlated with gene body methylation and enriched for genes encoding effector/secreted proteins
We next investigated if any of the heterozygous biallelic genes displayed ASE in any condition using our ONT cDNA data sets.
We first tested if there was an overall gene expression bias at the individual nuclear haplotype level. Both haplotypes displayed balanced expression without evidence of nuclear dominance (Supplemental Fig. S12); however, an expression bias for haplotype A was detected when considering only the heterozygous biallelic genes (Fig. 6B).
ASE is prevalent among secretome genes and correlates with gene methylation patterns. (A) The five ASE categories detected across six different transcript sampling conditions. Allele pairs displaying absolute log2 fold changes greater than two (Diff2 and Diff4) in at least one condition were defined as ASE in subsequent analysis. (B) Expression levels of alleles belonging to the two nuclear haplotypes A and B. Mann–Whitney U test: (***) P < 0.001. (C) Odds ratios and log10 P-values from Fisher's exact tests comparing the proportion of ASE genes among secretome genes and evolutionarily conserved BUSCOs. Each red dot represents a transcript sampling conditions as labeled. Dotted red and gray lines highlight cut-offs matching the null hypothesis of no difference between the two gene groups. (D) Distribution of the percentage of CpG methylation (sampled at UG) across ASE or non-ASE secretome gene bodies (here defined as start to stop codon) including ±2 kbp flanking regions. Solid lines represent mean percentage of CpG methylation; shaded areas represent 95% bootstrapping confidence intervals. Yellow inset shows the number of allele pairs included.
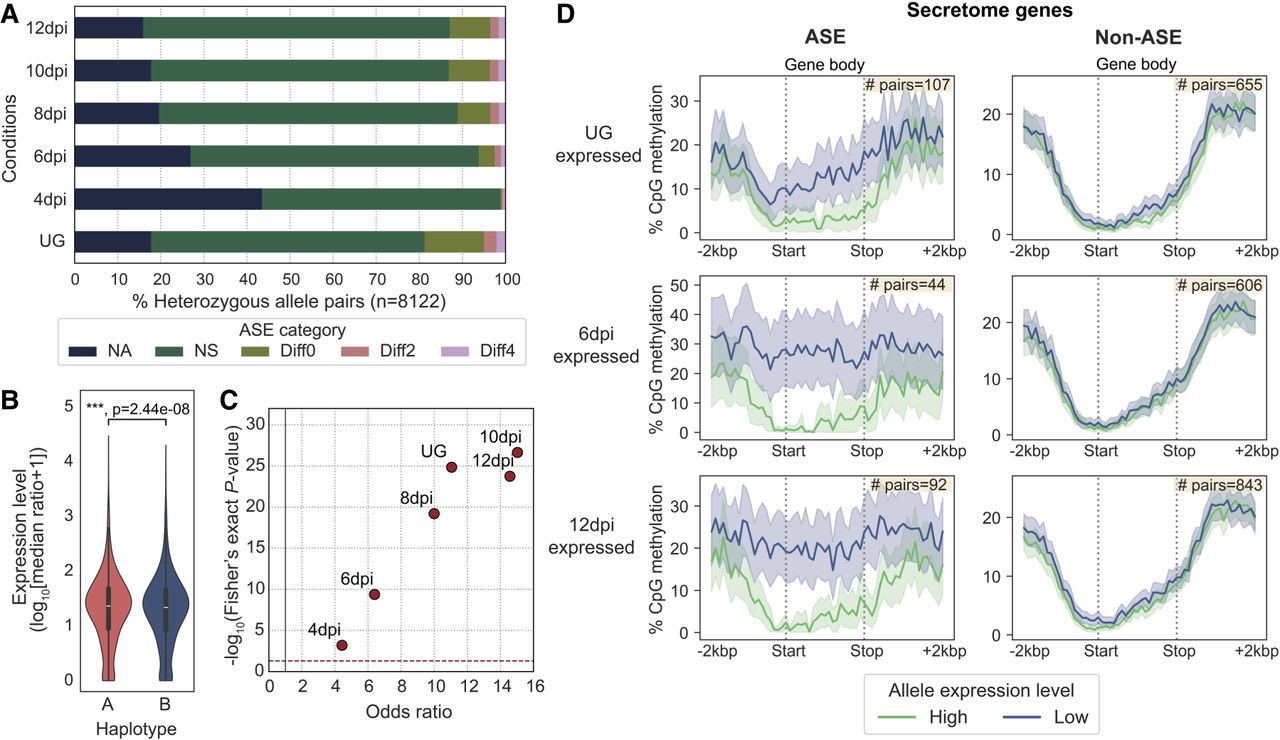
We compared the transcript abundance for each heterozygous allele pair to determine its ASE across all six tested conditions (Fig. 6A; Supplemental Table S12). We classified the ASE status using the following criteria (Shi et al. 2024): (1) no observed expression or no unambiguous transcript mapping at both alleles (NA), (2) no significant difference between alleles with false-discovery rate (FDR) adjusted P-value > 0.05 (NS), and (3) significant difference between alleles with adjusted P-value < 0.05, indicating differential ASE. The differential ASE pairs were further classified based on log2 fold change (LFC): weak ASE, |LFC| < 2 (Diff0); moderate ASE, 2 ≤ |LFC| < 4 (Diff2); and strong ASE, |LFC| ≥ 4 (Diff4). Although most heterozygous genes showed no evidence of ASE, a substantial proportion exhibited ASE in each condition. The UG stage had most ASE pairs (18.7%), followed by 8 to 12 dpi, which maintained stable ASE rates (11.1%–13.2%). Early infection at 4 dpi had the fewest (1.2%) ASE pairs, potentially owing to the low fungal biomass sampled. To reduce noise in the subsequent ASE analyses, we applied a |LFC| threshold of two to define the ASE set (Diff2 and Diff4), whereas all other categories were defined as non-ASE for the remainder of this study. Intersection analysis identified 678 allele pairs (8.3%) that were assigned to Diff2 or Diff4 ASE in at least one condition (Supplemental Fig. S13).
Given that pathogenicity-related effectors typically undergo diversifying selection (Stukenbrock and McDonald 2009; Sperschneider et al. 2014), we hypothesized that ASE might be an additional pathway to drive virulence dynamics. We therefore asked whether secretome genes, including putative effectors, were overrepresented in each condition-specific ASE versus non-ASE set compared with those of evolutionarily conserved BUSCOs. A Fisher's exact test (FDR < 5%) for every condition consistently showed significant ASE enrichment for secretome genes relative to BUSCOs with odds ratios greater than one (Fig. 6C; Supplemental Table S13, see contingency tables).
We explored several epigenetic and genetic factors that potentially underlie the ASE of secretome genes. No association was found between ASE and SVs (Fig. 5C), TE occupancy, and allelic sequence divergence (Supplemental Figs. S14, S15; Supplemental Note). We compared the CpG methylation density between secretome alleles along their gene bodies (start to stop codon) and 2 kbp flanking sequences to include proximal cis-regulatory elements such as promoters. Significant methylation differences were observed around the start codon between ASE alleles across all in planta infection conditions compared with the non-ASE alleles (Fig. 6D; Supplemental Fig. S16). Higher-expressed alleles were found to be hypomethylated at the start codon windows (1.1%–2.5% CpGs methylated), whereas those of their lower-expressed counterparts were frequently more heavily methylated (19.2%–36.1% CpGs methylated). Non-ASE secretome genes showed no methylation difference between alleles, regardless of their expression levels. These data suggest that DNA methylation imbalance may be involved in secretome ASE.
Discussion
Recent advances in long-read sequencing technologies have made T2T haplotype-phased genome assemblies the new gold standard for eukaryotes, including di- and heterokaryotic fungi such as rust fungi. Here, we report a fully nuclear-phased T2T genome assembly for Pst, the first reconstructed using high-accuracy ONT duplex sequencing. We show that our ONT-only T2T genome assembly is of comparable or superior quality to recently published PacBio HiFi-based haplotype-resolved T2T assemblies of other di- and heterokaryotic fungi (e.g., Sperschneider et al. 2023a; Henningsen et al. 2024; Wang et al. 2024). With the complete resolution of both nuclear haplotypes in our Pst assembly, we were able to uncover novel insights into the impact of dikaryotism on the genome biology of a long-term asexual clonal isolate of this fungus.
Our detailed analyses of Pst centromeres show that they adopt the classic Rabl configuration with clustering of heterochromatic centromeres (Xia et al. 2022; Torres et al. 2023). This arrangement allowed us to identify large regional centromeres from Hi-C contact hotspots that coincide with hypermethylated, gene-poor genomic signatures. Each centromere has a single potential kinetochore attachment site marked by a hypomethylated pocket known as the CDR (Logsdon and Eichler 2023; Logsdon et al. 2024; Mastrorosa et al. 2024), within otherwise fully CpG-methylated centromeres without signs of elevated AT content. Direct experimental evidence from CENP-A (CenH3) chromatin immunoprecipitation sequencing will be required to validate centromere localization. Our comparative inter-haplotype analysis revealed that Pst centromeres are highly variable in length and sequence, lacking characteristic motifs that define all centromeres. In most cases, the inferred kinetochore site appears to be consistently positioned in homologous chromosomes even considering highly divergent homologous centromeres, and where shifts occur, we did not detect an associated sequence presence/absence pattern. These findings point to the conclusion that formation of Pst centromeres is a sequence-independent process, consistent with observations in many other fungal regional centromeres (Roy and Sanyal 2011; Smith et al. 2012; Schotanus et al. 2015; Sperschneider et al. 2021; Cissé et al. 2024). Most but not all Pst centromeres are enriched for LTR retrotransposons, especially the Ty3/Gypsy superfamily. LTR-rich centromeres have also been reported for other pathogenic fungi with large regional type centromeres, such as the closely related stem rust fungus Puccinia graminis f. sp. tritici (Sperschneider et al. 2021), a human pathogenic yeast Cryptococcus neoformans (Yadav et al. 2018b), and an ascomycetous phytopathogen Verticillium dahliae (Seidl et al. 2020), but direct roles for LTR retrotransposons in centromere establishment remain unclear. In C. neoformans, the RNAi machinery and DNA methylation have been proposed as key epigenetic drivers for centromere identity via suppressing transposition and deleterious recombination among centromeric LTRs (Yadav et al. 2018b). However, the fact that the correlation with LTR enrichment does not hold in some Pst centromeres supports the idea that the LTR sequence itself does not define centromeres (Lynch et al. 2010; Guin et al. 2020). Rather, LTRs might be preferentially inserted owing to their high proliferative potential, which may, in turn, promote RNAi-directed silencing to reinforce centromere formation (Balzano and Giunta 2020). Further studies will be required to elucidate such links in Pst.
The dikaryotic configuration of Pst prompted us to explore its effect on intraspecific rDNA dynamics under strict clonality. Generally, rDNA tandem repeats are thought to undergo concerted evolution toward sequence homogenization via repeated homologous recombination, namely, unequal crossovers and gene conversion (Symonová 2019; Mullis et al. 2020; Garcia et al. 2024). Because Pst104E has genetically distinct nuclei, we hypothesized that its rDNA variants may persist or emerge within individual nuclei in the absence of meiotic exchange. Our results show that each nucleus of Pst104E carries a unique array with more than 200 repeats, predominated by an rDNA sequence subtype that harbors nucleus-specific variations within both IGS regions. A low proportion of these repeats have diversified through accumulating point mutations, and some appeared to have become fixed. We speculate that such nucleus-specific rDNA subtype homogeneity might be the consequence of compartmentalized concerted evolution owing to the individual inheritance of each nucleus over prolonged clonal history. The intra-array diversification, however, signifies a relaxation of concerted evolution, leading to incomplete homogenization within each nucleus (Xu et al. 2017; Wang et al. 2023). A possible explanation for this could be the reliance on limited nonmeiotic homologous recombination (e.g., intrachromosomal or between sister chromatids), which might less efficiently purge the newly spreading variants (Paloi et al. 2022). This parallels previous observations from homokaryotic (i.e., has genetically uniform nuclei) arbuscular mycorrhizal fungi strains (Serghi et al. 2021), which displayed extensive rDNA heterogeneity within each nucleus, consistent with their ancient clonality (Pawlowska and Taylor 2004; Lin et al. 2014). In the future, it will be interesting to survey intraspecific rDNA variations in rust isolates that have arisen via recent sexual recombination (Heitman et al. 2013; Wallen and Perlin 2018; Wang et al. 2022), in which we expect that divergent rDNA subtypes will be more evenly distributed among nuclei. Dikaryotism therefore presents an excellent opportunity for understanding the rates and dynamics of concerted evolution in fungi.
A fundamental question in genome biology is whether ASE could have functional consequences that lead to phenotypic variability (Cleary and Seoighe 2021; St. Pierre et al. 2022). Here, we show that ASE is pervasive in Pst and appears to be inversely related to gene body methylation, in which lower-expressed alleles display higher levels of CpG methylation. In Pst104E, ASE is overrepresented in secretome genes including putative effectors that could be involved in host pathogenesis. Therefore, ASE might present a novel transcriptomic regulatory mechanism to generate effector diversity beyond protein sequence variations (Chen et al. 2017; Salcedo et al. 2017; Ortiz et al. 2022). Expression-level polymorphisms between recognized and nonrecognized effector alleles may also explain virulence switching. For example, in P. graminis f. sp. tritici, a virulence allele of AvrSr27 was expressed at a much lower level than its avirulence allele counterpart, yet when overexpressed in planta, recognition took place (Upadhyaya et al. 2021). Similar observations have been recently made in a common rust disease of maize caused by Puccinia sorghi. A lowly expressed virulence allele of AvrRp1-D, differing by only one amino acid from its avirulence counterpart, became recognized by the cognate resistance gene Rp1-D after co-overexpression in planta (Kim et al. 2024). Nonrecognition can be therefore be caused by low expression rather than an inability of the host and pathogen proteins to interact. Future studies will inform whether the ASE of these P. graminis f. sp. tritici and P. sorghi effector genes is linked to changes in gene body methylation. The role of epigenetic regulation of Avr gene expression was previously highlighted in the soybean pathogen Phytophthora sojae, in which the natural silencing of the avirulence gene Avr3a resulted in gain-of-virulence (Qutob et al. 2013; Hale et al. 2023). Such epigenetics-mediated ASE may offer a reversible means to selectively silence or “archive” avirulence alleles to escape immune recognition while retaining the unmutated gene.
One important limitation of our study is that our DNA methylation data only reflects dormancy. A previous study in Zymoseptoria tritici (Meile et al. 2020) demonstrated that chromatin remodeling via histone modifications, which are highly correlated with DNA methylation in fungi (Rose and Klose 2014; He et al. 2020), can occur during host pathogenesis to derepress effector expression. To extend our observations, we will need DNA methylation and histone modification data sampled from Pst undergoing pathogenesis to determine if chromatin dynamism underpins ASE in planta.
Flor's classic “gene-for-gene” hypothesis (Flor 1942) that has shaped our understanding of plant resistance to pathogens since the 1940s might be an oversimplification, and we may have to consider that differences in effector gene expression underlie disease outcomes in the field.
Methods
Methods for Pst104E DNA and RNA extraction, ONT long-read genome and transcriptome sequencing, and Hi-C library sequencing are detailed in the Supplemental Methods.
Genome assembly and quality evaluation
Filtered duplex (>10 kbp; Q30; 32×/haplotype) and simplex (>40 kbp; Q10; 117×/haplotype) reads were assembled using Verkko v1.3.1 (Rautiainen et al. 2023). The assembly was scaffolded with Hi-C using Juicer v2.0 (Durand et al. 2016b) and 3D-DNA v180114 (Dudchenko et al. 2017), followed by manual curation (Supplemental Methods). Each haplotype was quality-assessed using assembly statistics, BUSCO v5.5.0 (basidiomycota_odb10) (Manni et al. 2021), Merqury v1.3 (Rhie et al. 2020), and LAI (Ou et al. 2018; Ou and Jiang 2018). CRAQ v1.0.9 (Li et al. 2023b) was launched to compute R/S-AQI inferred from regional and structural errors from clipped alignments that may indicate misjoins. To evaluate phasing quality, HiC-Pro v3.1.0 (Servant et al. 2015) was used to generate contact matrices from Hi-C alignments (MAPQ ≥ 20), which were analyzed using scripts from GitHub (https://github.com/RunpengLuo/HiC-Analysis) to quantify cis- and trans-chromosome contacts.
TE and gene annotations
TE and gene annotations were performed for each haplotype separately. We predicted and annotated TEs using the REPET pipeline v3.0 (Quesneville et al. 2005; Flutre et al. 2011). Prior to gene annotation, we filtered host RNA from all transcriptomic data sets by mapping them against the Pst104E assembly with minimap2 v2.26 (Li 2018) and retaining only the mappable reads. We then independently processed and assembled the Illumina RNA-seq and the ONT cDNA data sets to generate transcript evidence. For ONT cDNA, reads were trimmed with Porechop_ABI v0.5.0 (Bonenfant et al. 2022), aligned to the dikaryotic assembly with minimap2 in splice-aware mode (-ax splice -ub -G 3000 ‐‐secondary=no), and partitioned into haplotype sets. To identify transcript structures from noisy long reads reference-guided and annotation-free, we employed two long-read-tailored tools: StringTie2 (-L -s2 -m50) (Kovaka et al. 2019) for better single-exon transcript discovery and ESPRESSO v1.4.0 (ESPRESSO_S.pl -Q0) (Gao et al. 2023) for improved splice site detection. Transcript annotations were merged across all samples using StringTie2 (‐‐merge). Gene predictions and functional annotations were performed using funannotate v1.8.15 (https://github.com/nextgenusfs/funannotate). For details, see the Supplemental Methods.
Differential expression analysis
Transcript abundance was quantified from ONT cDNA spliced alignments using Bambu v3.4.1 (Chen et al. 2023) with the isoform discovery mode disabled. The resulting count matrices were imported to DESeq2 v1.38.3 (Love et al. 2014) for analysis. PCA was performed on variance-stabilizing transformed read counts to visualize sample clustering. DESeq2 was executed on default settings to identify genes differentially expressed at host infection conditions relative to UG. We considered genes with an FDR-adjusted P-value < 0.05 and LFC ≥ 2 to be upregulated. Secretome genes upregulated at 4, 6, or 8 dpi were identified as our preliminary Avr effector candidates.
Centromere inference and analysis
Centromere locations were determined based on strong inter-chromosomal interaction signals on Juicebox v2.17.00 Hi-C heatmap (Durand et al. 2016a) and confirmed by analyzing ONT-derived DNA methylation data (Supplemental Methods).
CDRs were located via visually selecting the largest hypomethylated region as this single pattern consistently appeared throughout all the inferred centromeres. Relative CDR position was calculated by dividing its midpoint coordinate by centromere length and then compared between haplotypes to detect CDR shifts. High-resolution alignment dotplots were generated in Gepard v2.1 to investigate the sequence composition at shifted CDRs (Krumsiek et al. 2007).
Centromeric TE enrichment was analyzed with permutation tests using our custom Python script. The rationale behind was to randomly reshuffle locations of target features (e.g., TEs) throughout a given chromosome or genome (5000 times) to remove their potential biological association with a region of interest (e.g., centromere), creating a null distribution of test statistics (e.g., TE coverage difference between centromere vs. noncentromere), which enabled a two-tailed hypothesis test for feature enrichment or depletion. P-values were defined as the proportion of permuted values equal to or more extreme than the observed value (<5% FDR).
rDNA analysis
The canonical rDNA unit was defined by aligning reference ITS, 18S, and 5S of Puccinia species retrieved from databases including Gold Standard (Eenjes et al. 2022), EukRibo (Berney et al. 2022), and 5SrRNAdb (Szymanski et al. 2016) to distinguish conserved and variable elements. Long rRNA reads were aligned to confirm the transcribed units. Analyses of rDNA subtype variations and copy numbers were conducted on duplex and Illumina read alignments (Supplemental Methods).
The nuclear association of the two dominant rDNA subtypes was tested by analyzing rDNA Hi-C reads with a k-mer approach. Unique 31-mers of each subtype were identified using UniqueKMER (Chen et al. 2021). Subtype-specific 31-mers, along with their reverse complements, were used to tag rDNA Hi-C reads, whose paired Hi-C mates were fetched from the corresponding R1/R2 read file. Mates were mapped to the dikaryotic assembly with bwa-mem2 (Vasimuddin et al. 2019). Mapping locations and MAPQ scores were extracted with BEDTools bamtobed (Quinlan and Hall 2010) for plotting. This was repeated on subtype-nonspecific 31-mers as control. Note that if a read is tagged by both subtype-specific and unspecific k-mers, it is defined as subtype specific.
Synteny and SV detection
Whole-genome alignment between haplotypes A and B was conducted with NUCmer (‐‐maxmatch -l 200 -b 500 -c 500). Only alignment blocks with >90% identity were retained. SyRI (Goel et al. 2019) was used to annotate SVs, syntenic and unaligned regions and then visualized with plotsr (Goel and Schneeberger 2022). To analyze features within and nearby SVs, BEDTools slop was used to extend SV coordinates by 2 kbp in both directions. Enrichment or depletion for genomic features, including TEs, genes, and subsets such as secretome and ASE-only genes, was statistically assessed through two-tailed permutation tests, as described above.
Identification of hemizygous and heterozygous genes
Protein sequences of all annotated genes were analyzed using Proteinortho v6.3.1 (-synteny) (Lechner et al. 2011) to detect homologs between haplotypes. Genes lacking a hit on the alternative haplotype were considered hemizygous candidates. These were further filtered via reciprocal BLASTP (>70% identity; >70% query/subject coverage) to ensure the absence of alleles. One-to-one gene pairs with divergence values greater than zero (computed from codon-aware alignments generated using script “dN_dS_Pst134E.ipynb”) (Luo et al. 2024; https://github.com/ZhenyanLuo/codes-used-for-mating-type) were defined to be heterozygous biallelic.
ASE analysis
We reformatted the Bambu gene-level read count matrices to test differential allele expression at heterozygous biallelic genes per condition using DESeq2 (Supplemental Methods). Allele pairs were grouped into different ASE status, as detailed in the Results. To assess nuclear dominance, allele read counts were normalized using DESeq2's median-of-ratio method and transformed into expression levels as log10(median of ratio + 1) and then averaged across replicates for haplotype comparisons. Overrepresentation of secretome genes in the ASE set (Diff2 and Diff4) relative to BUSCOs was evaluated via a two-sided Fisher's exact test on a 2 × 2 contingency table constructed per condition (<5% FDR). Methylation differences between ASE and non-ASE secretome alleles at the gene body and flanking regions were analyzed using a custom Python script (Supplemental Methods).
Data access
All raw sequencing data generated in this study have been submitted to the NCBI BioProject database (https://www.ncbi.nlm.nih.gov/bioproject/) under accession number PRJNA1195871. All custom scripts for analyses and figures are available at GitHub (https://github.com/ritatam/Pst104EGenomeAnalysis) and as Supplemental Scripts. Genome assembly, rDNA sequences, and gene and TE annotations are available at Zenodo (https://doi.org/10.5281/zenodo.14885411) and as Supplemental Material.
Competing interest statement
The authors declare no competing interests.
Acknowledgments
R.T. was supported by a Grains Research and Development Corporation Graduate Research Scholarship. This work was supported by an Australian Research Council Future fellowship to B.S. (FT180100024) and a Discovery Project grant (DP230100941) to J.P.R. and B.S. This work was supported by computational resources provided by the Australian Government through the National Computational Infrastructure (NCI) under the ANU Merit Allocation Scheme. We acknowledge the contribution of the Plant Pathogen ‘Omics Initiative consortium in the generation of data used in this publication. The Initiative is supported by funding from Bioplatforms Australia, enabled by the Commonwealth Government National Collaborative Research Infrastructure Strategy (NCRIS).
Author contributions: R.T., B.S., and J.P.R. designed the project. R.T. led the project and performed most of the formal analyses and visualization. B.S. and J.P.R. supervised the project. R.T., B.S., J.P.R., and M.M. acquired funding. M.M. conducted RNA extraction and ONT cDNA sequencing. R.L. analyzed Hi-C data for evaluating assembly phasing quality. Z.L. identified mating type loci and shared scripts for allele pairing. B.S., A.J., and S.P. contributed to experiments. R.T., M.M., J.P.R., and B.S. wrote the manuscript. All authors have reviewed the final manuscript.
Notes
[1] Supplementary material [Supplemental material is available for this article.]
[2] Article published online before print. Article, supplemental material, and publication date are at https://www.genome.org/cgi/doi/10.1101/gr.280359.124.
[3] Freely available online through the Genome Research Open Access option.
References
- ↵Akiyoshi B. 2019. Evolution: a mosaic-type centromere in an early-diverging fungus. Curr Biol 29: R1184–R1186. 10.1016/j.cub.2019.09.042
- ↵Anderson JB, Kohn LM. 2014. Dikaryons, diploids, and evolution. In Sex in fungi (ed. Heitman J, ), pp. 333–348. ASM Press, Washington, DC.
- ↵Balzano E, Giunta S. 2020. Centromeres under pressure: evolutionary innovation in conflict with conserved function. Genes (Basel) 11: 912. 10.3390/genes11080912
- ↵Belser C, Baurens F-C, Noel B, Martin G, Cruaud C, Istace B, Yahiaoui N, Labadie K, Hřibová E, Doležel J, 2021. Telomere-to-telomere gapless chromosomes of banana using nanopore sequencing. Commun Biol 4: 1047. 10.1038/s42003-021-02559-3
- ↵Berney C, Henry N, Mahé F, Richter DJ, de Vargas C. 2022. Eukribo: a manually curated eukaryotic 18S rDNA reference database to facilitate identification of new diversity. bioRxiv 10.1101/2022.11.03.515105
- ↵Bodor DL, Mata JF, Sergeev M, David AF, Salimian KJ, Panchenko T, Cleveland DW, Black BE, Shah JV, Jansen LE. 2014. The quantitative architecture of centromeric chromatin. eLife 3: e02137. 10.7554/eLife.02137
- ↵Bonenfant Q, Noé L, Touzet H. 2022. Porechop_ABI: discovering unknown adapters in Oxford Nanopore Technology sequencing reads for downstream trimming. Bioinform Adv 3: vbac085. 10.1093/bioadv/vbac085
- ↵Castanera R, Borgognone A, Pisabarro AG, Ramírez L. 2017. Biology, dynamics, and applications of transposable elements in basidiomycete fungi. Appl Microbiol Biotechnol 101: 1337–1350. 10.1007/s00253-017-8097-8
- ↵Chen J, Upadhyaya NM, Ortiz D, Sperschneider J, Li F, Bouton C, Breen S, Dong C, Xu B, Zhang X, 2017. Loss of AvrSr50 by somatic exchange in stem rust leads to virulence for Sr50 resistance in wheat. Science 358: 1607–1610. 10.1126/science.aao4810
- ↵Chen S, He C, Li Y, Li Z, Melançon CEIII. 2021. A computational toolset for rapid identification of SARS-CoV-2, other viruses and microorganisms from sequencing data. Brief Bioinform 22: 924–935. 10.1093/bib/bbaa231
- ↵Chen Y, Sim A, Wan YK, Yeo K, Lee JJX, Ling MH, Love MI, Göke J. 2023. Context-aware transcript quantification from long-read RNA-seq data with Bambu. Nat Methods 20: 1187–1195. 10.1038/s41592-023-01908-w
- ↵Cissé OH, Curran SJ, Folco HD, Liu Y, Bishop L, Wang H, Fischer ER, Davis AS, Combs C, Thapar S, 2024. Regional centromere configuration in the fungal pathogens of the Pneumocystis genus. mBio 15: e03185-23. 10.1128/mbio.03185-23
- ↵Cleary S, Seoighe C. 2021. Perspectives on allele-specific expression. Annu Rev Biomed Data Sci 4: 101–122. 10.1146/annurev-biodatasci-021621-122219
- ↵Corre E, Morin E, Duplessis S, Lorrain C. 2025. Ancestral and recent bursts of transposition shaped the massive genomes of plant pathogenic rust fungi. bioRxiv 10.1101/2025.01.10.632365
- ↵Cuomo CA, Bakkeren G, Khalil HB, Panwar V, Joly D, Linning R, Sakthikumar S, Song X, Adiconis X, Fan L, 2017. Comparative analysis highlights variable genome content of wheat rusts and divergence of the mating loci. G3 (Bethesda) 7: 361–376. 10.1534/g3.116.032797
- ↵Dobon A, Bunting DCE, Cabrera-Quio LE, Uauy C, Saunders DGO. 2016. The host-pathogen interaction between wheat and yellow rust induces temporally coordinated waves of gene expression. BMC Genomics 17: 380. 10.1186/s12864-016-2684-4
- ↵Du Z, Peng Y, Zhang G, Chen L, Jiang S, Kang Z, Zhao J. 2023. Direct evidence demonstrates that Puccinia striiformis f. sp. tritici infects susceptible barberry to complete sexual cycle in autumn. Plant Dis 107: 771–783. 10.1094/PDIS-08-22-1750-RE
- ↵Dudchenko O, Batra SS, Omer AD, Nyquist SK, Hoeger M, Durand NC, Shamim MS, Machol I, Lander ES, Aiden AP, 2017. De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science 356: 92–95. 10.1126/science.aal3327
- ↵Durand NC, Robinson JT, Shamim MS, Machol I, Mesirov JP, Lander ES, Aiden EL. 2016a. Juicebox provides a visualization system for Hi-C contact maps with unlimited zoom. Cell Syst 3: 99–101. 10.1016/j.cels.2015.07.012
- ↵Durand NC, Shamim MS, Machol I, Rao SSP, Huntley MH, Lander ES, Aiden EL. 2016b. Juicer provides a one-click system for analyzing loop-resolution Hi-C experiments. Cell Syst 3: 95–98. 10.1016/j.cels.2016.07.002
- ↵Eenjes T, Hu Y, Irinyi L, Hoang MTV, Smith LM, Linde CC, Milgate AW, Meyer W, Stone EA, Rathjen JP, 2022. Linked machine learning classifiers improve species classification of fungi when using error-prone long-reads on extended metabarcodes. bioRxiv 10.1101/2021.05.01.442223
- ↵Ferguson S, Jones A, Murray K, Andrew R, Schwessinger B, Borevitz J. 2024. Plant genome evolution in the genus eucalyptus is driven by structural rearrangements that promote sequence divergence. Genome Res 34: 606–619. 10.1101/gr.277999.123
- ↵Flor HH. 1942. Inheritance of pathogenicity in Melampsora lini. Phytopathol 32: 653–669.
- ↵Flutre T, Duprat E, Feuillet C, Quesneville H. 2011. Considering transposable element diversification in de novo annotation approaches. PLoS One 6: e16526. 10.1371/journal.pone.0016526
- ↵Fultz D, McKinlay A, Enganti R, Pikaard CS. 2023. Sequence and epigenetic landscapes of active and silent nucleolus organizer regions in Arabidopsis. Sci Adv 9: eadj4509. 10.1126/sciadv.adj4509
- ↵Gao Y, Wang F, Wang R, Kutschera E, Xu Y, Xie S, Wang Y, Kadash-Edmondson KE, Lin L, Xing Y. 2023. ESPRESSO: robust discovery and quantification of transcript isoforms from error-prone long-read RNA-seq data. Sci Adv 9: eabq5072. 10.1126/sciadv.abq5072
- ↵Garcia S, Kovarik A, Maiwald S, Mann L, Schmidt N, Pascual-Díaz JP, Vitales D, Weber B, Heitkam T. 2024. The dynamic interplay between ribosomal DNA and transposable elements: a perspective from genomics and cytogenetics. Mol Biol Evol 41: msae025. 10.1093/molbev/msae025
- ↵Gehrmann T, Pelkmans JF, Ohm RA, Vos AM, Sonnenberg ASM, Baars JJP, Wösten HAB, Reinders MJT, Abeel T. 2018. Nucleus-specific expression in the multinuclear mushroom-forming fungus Agaricus bisporus reveals different nuclear regulatory programs. Proc Natl Acad Sci 115: 4429–4434. 10.1073/pnas.1721381115
- ↵Gluck-Thaler E, Ralston T, Konkel Z, Ocampos CG, Ganeshan VD, Dorrance AE, Niblack TL, Wood CW, Slot JC, Lopez-Nicora HD, 2022. Giant starship elements mobilize accessory genes in fungal genomes. Mol Biol Evol 39: msac109. 10.1093/molbev/msac109
- ↵Goel M, Schneeberger K. 2022. plotsr: visualizing structural similarities and rearrangements between multiple genomes. Bioinformatics 38: 2922–2926. 10.1093/bioinformatics/btac196
- ↵Goel M, Sun H, Jiao W-B, Schneeberger K. 2019. SyRI: finding genomic rearrangements and local sequence differences from whole-genome assemblies. Genome Biol 20: 277. 10.1186/s13059-019-1911-0
- ↵Grünberger F, Ferreira-Cerca S, Grohmann D. 2022. Nanopore sequencing of RNA and cDNA molecules in Escherichia coli. RNA 28: 400–417. 10.1261/rna.078937.121
- ↵Guin K, Sreekumar L, Sanyal K. 2020. Implications of the evolutionary trajectory of centromeres in the fungal kingdom. Annu Rev Microbiol 74: 835–853. 10.1146/annurev-micro-011720-122512
- ↵Hale B, Brown E, Wijeratne A. 2023. An updated assessment of the soybean–Phytophthora sojae pathosystem. Plant Pathol 72: 843–860. 10.1111/ppa.13713
- ↵Hartmann FE. 2022. Using structural variants to understand the ecological and evolutionary dynamics of fungal plant pathogens. New Phytol 234: 43–49. 10.1111/nph.17907
- ↵He C, Zhang Z, Li B, Tian S. 2020. The pattern and function of DNA methylation in fungal plant pathogens. Microorganisms 8: 227. 10.3390/microorganisms8020227
- ↵Heitman J, Sun S, James TY. 2013. Evolution of fungal sexual reproduction. Mycologia 105: 1–27. 10.3852/12-253
- ↵Henningsen EC, Lewis D, Nazareno ES, Mangelson H, Sanchez M, Langford K, Huang YF, Steffenson BJ, Boesen B, Kianian SF, 2024. A high-resolution haplotype collection uncovers somatic hybridization, recombination and intercontinental movement in oat crown rust. PLoS Genet 20: e1011493. 10.1371/journal.pgen.1011493
- ↵Hu G, Grover CE, Arick MA, Liu M, Peterson DG, Wendel JF. 2021. Homoeologous gene expression and co-expression network analyses and evolutionary inference in allopolyploids. Brief Bioinform 22: 1819–1835. 10.1093/bib/bbaa035
- ↵James TY, Stajich JE, Hittinger CT, Rokas A. 2020. Toward a fully resolved fungal tree of life. Annu Rev Microbiol 74: 291–313. 10.1146/annurev-micro-022020-051835
- ↵Kim S-B, Kim K-T, In S, Jaiswal N, Lee G-W, Jung S, Rogers A, Gómez-Trejo LF, Gautam S, Helm M, 2024. Use of the Puccinia sorghi haustorial transcriptome to identify and characterize AvrRp1-D recognized by the maize Rp1-D resistance protein. PLoS Pathog 20: e1012662. 10.1371/journal.ppat.1012662
- ↵Kovaka S, Zimin AV, Pertea GM, Razaghi R, Salzberg SL, Pertea M. 2019. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol 20: 278. 10.1186/s13059-019-1910-1
- ↵Krumsiek J, Arnold R, Rattei T. 2007. Gepard: a rapid and sensitive tool for creating dotplots on genome scale. Bioinformatics 23: 1026–1028. 10.1093/bioinformatics/btm039
- ↵Lechner M, Findeiß S, Steiner L, Marz M, Stadler PF, Prohaska SJ. 2011. Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 12: 124. 10.1186/1471-2105-12-124
- ↵Li H. 2018. Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34: 3094–3100. 10.1093/bioinformatics/bty191
- ↵Li F, Upadhyaya NM, Sperschneider J, Matny O, Nguyen-Phuc H, Mago R, Raley C, Miller ME, Silverstein KAT, Henningsen E, 2019. Emergence of the Ug99 lineage of the wheat stem rust pathogen through somatic hybridisation. Nat Commun 10: 5068. 10.1038/s41467-019-12927-7
- ↵Li C, Qiao L, Lu Y, Xing G, Wang X, Zhang G, Qian H, Shen Y, Zhang Y, Yao W, 2023a. Gapless genome assembly of Puccinia triticina provides insights into chromosome evolution in Pucciniales. Microbiol Spectr 11: e02828-22. 10.1128/spectrum.02828-22
- ↵Li K, Xu P, Wang J, Yi X, Jiao Y. 2023b. Identification of errors in draft genome assemblies at single-nucleotide resolution for quality assessment and improvement. Nat Commun 14: 6556. 10.1038/s41467-023-42336-w
- ↵Lin K, Limpens E, Zhang Z, Ivanov S, Saunders DGO, Mu D, Pang E, Cao H, Cha H, Lin T, 2014. Single nucleus genome sequencing reveals high similarity among nuclei of an endomycorrhizal fungus. PLoS Genet 10: e1004078. 10.1371/journal.pgen.1004078
- ↵Logsdon GA, Eichler EE. 2023. The dynamic structure and rapid evolution of human centromeric satellite DNA. Genes (Basel) 14: 92. 10.3390/genes14010092
- ↵Logsdon GA, Rozanski AN, Ryabov F, Potapova T, Shepelev VA, Catacchio CR, Porubsky D, Mao Y, Yoo D, Rautiainen M, 2024. The variation and evolution of complete human centromeres. Nature 629: 136–145. 10.1038/s41586-024-07278-3
- ↵Love MI, Huber W, Anders S. 2014. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15: 550. 10.1186/s13059-014-0550-8
- ↵Luo Z, McTaggart A, Schwessinger B. 2024. Genome biology and evolution of mating-type loci in four cereal rust fungi. PLoS Genet 20: e1011207. 10.1371/journal.pgen.1011207
- ↵Lynch DB, Logue ME, Butler G, Wolfe KH. 2010. Chromosomal G + C content evolution in yeasts: systematic interspecies differences, and GC-poor troughs at centromeres. Genome Biol Evol 2: 572–583. 10.1093/gbe/evq042
- ↵Manni M, Berkeley MR, Seppey M, Simão FA, Zdobnov EM. 2021. BUSCO update: novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol Biol Evol 38: 4647–4654. 10.1093/molbev/msab199
- ↵Mastrorosa FK, Rozanski AN, Harvey WT, Knuth J, Garcia G, Munson KM, Hoekzema K, Logsdon GA, Eichler EE. 2024. Complete chromosome 21 centromere sequences from a down syndrome family reveal size asymmetry and differences in kinetochore attachment. bioRxiv 10.1101/2024.02.25.581464
- ↵Meile L, Peter J, Puccetti G, Alassimone J, McDonald BA, Sánchez-Vallet A. 2020. Chromatin dynamics contribute to the spatiotemporal expression pattern of virulence genes in a fungal plant pathogen. mBio 11: e02343-20. 10.1128/mBio.02343-20
- ↵Muller H, Gil J, Drinnenberg IA. 2019. The impact of centromeres on spatial genome architecture. Trends Genet 35: 565–578. 10.1016/j.tig.2019.05.003
- ↵Mullis A, Lu Z, Zhan Y, Wang T-Y, Rodriguez J, Rajeh A, Chatrath A, Lin Z. 2020. Parallel concerted evolution of ribosomal protein genes in fungi and its adaptive significance. Mol Biol Evol 37: 455–468. 10.1093/molbev/msz229
- ↵Narayanan A, Reza MH, Sanyal K. 2024. Behind the scenes: centromere-driven genomic innovations in fungal pathogens. PLoS Pathog 20: e1012080. 10.1371/journal.ppat.1012080
- ↵Nurk S, Koren S, Rhie A, Rautiainen M, Bzikadze AV, Mikheenko A, Vollger MR, Altemose N, Uralsky L, Gershman A, 2022. The complete sequence of a human genome. Science 376: 44–53. 10.1126/science.abj6987
- ↵Ortiz D, Chen J, Outram MA, Saur IML, Upadhyaya NM, Mago R, Ericsson DJ, Cesari S, Chen C, Williams SJ, 2022. The stem rust effector protein AvrSr50 escapes Sr50 recognition by a substitution in a single surface-exposed residue. New Phytol 234: 592–606. 10.1111/nph.18011
- ↵Ou S, Jiang N. 2018. LTR_retriever: a highly accurate and sensitive program for identification of long terminal repeat retrotransposons. Plant Physiol 176: 1410–1422. 10.1104/pp.17.01310
- ↵Ou S, Chen J, Jiang N. 2018. Assessing genome assembly quality using the LTR assembly index (LAI). Nucleic Acids Res 46: e126. 10.1093/nar/gky730
- ↵Paloi S, Luangsa-ard JJ, Mhuantong W, Stadler M, Kobmoo N. 2022. Intragenomic variation in nuclear ribosomal markers and its implication in species delimitation, identification and barcoding in fungi. Fungal Biol Rev 42: 1–33. 10.1016/j.fbr.2022.04.002
- ↵Pardo-Palacios FJ, Wang D, Reese F, Diekhans M, Carbonell-Sala S, Williams B, Loveland JE, De María M, Adams MS, Balderrama-Gutierrez G, 2024. Systematic assessment of long-read RNA-seq methods for transcript identification and quantification. Nat Methods 21: 1349–1363. 10.1038/s41592-024-02298-3
- ↵Pawlowska TE, Taylor JW. 2004. Organization of genetic variation in individuals of arbuscular mycorrhizal fungi. Nature 427: 733–737. 10.1038/nature02290
- ↵Quesneville H, Bergman CM, Andrieu O, Autard D, Nouaud D, Ashburner M, Anxolabehere D. 2005. Combined evidence annotation of transposable elements in genome sequences. PLoS Comput Biol 1: e22. 10.1371/journal.pcbi.0010022
- ↵Quinlan AR, Hall IM. 2010. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26: 841–842. 10.1093/bioinformatics/btq033
- ↵Qutob D, Patrick Chapman B, Gijzen M. 2013. Transgenerational gene silencing causes gain of virulence in a plant pathogen. Nat Commun 4: 1349. 10.1038/ncomms2354
- ↵Ramírez Nasto L, Pérez Garrido MG, Castanera Andrés R, Santoyo Santos F, Pisabarro de Lucas G. 2011. Basidiomycetes telomeres: a bioinformatics approach. In Bioinformatics: Trends and Methodologies (ed. Badria FA), pp. 393–424. IntechOpen, London. https://academica-e.unavarra.es/handle/2454/35292
- ↵Rautiainen M, Nurk S, Walenz BP, Logsdon GA, Porubsky D, Rhie A, Eichler EE, Phillippy AM, Koren S. 2023. Telomere-to-telomere assembly of diploid chromosomes with Verkko. Nat Biotechnol 41: 1474–1482. 10.1038/s41587-023-01662-6
- ↵Rhie A, Walenz BP, Koren S, Phillippy AM. 2020. Merqury: reference-free quality, completeness, and phasing assessment for genome assemblies. Genome Biol 21: 245. 10.1186/s13059-020-02134-9
- ↵Rodriguez-Algaba J, Hovmøller MS, Schulz P, Hansen JG, Lezáun JA, Joaquim J, Randazzo B, Czembor P, Zemeca L, Slikova S, 2022. Stem rust on barberry species in Europe: host specificities and genetic diversity. Front Genet 13: 988031. 10.3389/fgene.2022.988031
- ↵Rose NR, Klose RJ. 2014. Understanding the relationship between DNA methylation and histone lysine methylation. Biochim Biophys Acta 1839: 1362–1372. 10.1016/j.bbagrm.2014.02.007
- ↵Roy B, Sanyal K. 2011. Diversity in requirement of genetic and epigenetic factors for centromere function in fungi. Eukaryot Cell 10: 1384–1395. 10.1128/EC.05165-11
- ↵Salcedo A, Rutter W, Wang S, Akhunova A, Bolus S, Chao S, Anderson N, De Soto MF, Rouse M, Szabo L, 2017. Variation in the AvrSr35 gene determines Sr35 resistance against wheat stem rust race Ug99. Science 358: 1604–1606. 10.1126/science.aao7294
- ↵Sankaranarayanan SR, Ianiri G, Coelho MA, Reza MH, Thimmappa BC, Ganguly P, Vadnala RN, Sun S, Siddharthan R, Tellgren-Roth C, 2020. Loss of centromere function drives karyotype evolution in closely related Malassezia species. eLife 9: e53944. 10.7554/eLife.53944
- ↵Schmidt-Dannert C. 2016. Biocatalytic portfolio of Basidiomycota. Curr Opin Chem Biol 31: 40–49. 10.1016/j.cbpa.2016.01.002
- ↵Schotanus K, Soyer JL, Connolly LR, Grandaubert J, Happel P, Smith KM, Freitag M, Stukenbrock EH. 2015. Histone modifications rather than the novel regional centromeres of Zymoseptoria tritici distinguish core and accessory chromosomes. Epigenetics Chromatin 8: 41. 10.1186/s13072-015-0033-5
- ↵Schwessinger B, Sperschneider J, Cuddy WS, Garnica DP, Miller ME, Taylor JM, Dodds PN, Figueroa M, Park RF, Rathjen JP. 2018. A near-complete haplotype-phased genome of the dikaryotic wheat stripe rust fungus Puccinia striiformis f. sp. tritici reveals high interhaplotype diversity. mBio 9: e02275-17. 10.1128/mBio.02275-17
- ↵Schwessinger B, Chen Y-J, Tien R, Vogt JK, Sperschneider J, Nagar R, McMullan M, Sicheritz-Ponten T, Sørensen CK, Hovmøller MS, 2020. Distinct life histories impact dikaryotic genome evolution in the rust fungus Puccinia striiformis causing stripe rust in wheat. Genome Biol Evol 12: 597–617. 10.1093/gbe/evaa071
- ↵Schwessinger B, Jones A, Albekaa M, Hu Y, Mackenzie A, Tam R, Nagar R, Milgate A, Rathjen JP, Periyannan S. 2022. A chromosome scale assembly of an Australian Puccinia striiformis f. sp. tritici isolate of the PstS1 lineage. Mol Plant-Microbe Interactions 35: 293–296. 10.1094/MPMI-09-21-0236-A
- ↵Seidl MF, Kramer HM, Cook DE, Fiorin GL, Van Den Berg GCM, Faino L, Thomma BPHJ. 2020. Repetitive elements contribute to the diversity and evolution of centromeres in the fungal genus verticillium. mBio 11: e01714-20. 10.1128/mBio.01714-20
- ↵Serghi EU, Kokkoris V, Cornell C, Dettman J, Stefani F, Corradi N. 2021. Homo- and dikaryons of the arbuscular mycorrhizal fungus Rhizophagus irregularis differ in life history strategy. Front Plant Sci 12: 715377. 10.3389/fpls.2021.715377
- ↵Servant N, Varoquaux N, Lajoie BR, Viara E, Chen C-J, Vert J-P, Heard E, Dekker J, Barillot E. 2015. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biol 16: 259. 10.1186/s13059-015-0831-x
- ↵Sessegolo C, Cruaud C, Da Silva C, Cologne A, Dubarry M, Derrien T, Lacroix V, Aury J-M. 2019. Transcriptome profiling of mouse samples using nanopore sequencing of cDNA and RNA molecules. Sci Rep 9: 14908. 10.1038/s41598-019-51470-9
- ↵Shao L, Xing F, Xu C, Zhang Q, Che J, Wang X, Song J, Li X, Xiao J, Chen L-L, 2019. Patterns of genome-wide allele-specific expression in hybrid rice and the implications on the genetic basis of heterosis. Proc Natl Acad Sci 116: 5653–5658. 10.1073/pnas.1820513116
- ↵Shi T-L, Jia K-H, Bao Y-T, Nie S, Tian X-C, Yan X-M, Chen Z-Y, Li Z-C, Zhao S-W, Ma H-Y, 2024. High-quality genome assembly enables prediction of allele-specific gene expression in hybrid poplar. Plant Physiol 195: 652–670. 10.1093/plphys/kiae078
- ↵Smith KM, Galazka JM, Phatale PA, Connolly LR, Freitag M. 2012. Centromeres of filamentous fungi. Chromosome Res 20: 635–656. 10.1007/s10577-012-9290-3
- ↵Sperschneider J, Ying H, Dodds PN, Gardiner DM, Upadhyaya NM, Singh KB, Manners JM, Taylor JM. 2014. Diversifying selection in the wheat stem rust fungus acts predominantly on pathogen-associated gene families and reveals candidate effectors. Front Plant Sci 5: 372. 10.3389/fpls.2014.00372
- ↵Sperschneider J, Jones AW, Nasim J, Xu B, Jacques S, Zhong C, Upadhyaya NM, Mago R, Hu Y, Figueroa M, 2021. The stem rust fungus Puccinia graminis f. sp. tritici induces centromeric small RNAs during late infection that are associated with genome-wide DNA methylation. BMC Biol 19: 203. 10.1186/s12915-021-01123-z
- ↵Sperschneider J, Hewitt T, Lewis DC, Periyannan S, Milgate AW, Hickey LT, Mago R, Dodds PN, Figueroa M. 2023a. Nuclear exchange generates population diversity in the wheat leaf rust pathogen Puccinia triticina. Nat Microbiol 8: 2130–2141. 10.1038/s41564-023-01494-9
- ↵Sperschneider J, Yildirir G, Rizzi YS, Malar CM, Mayrand Nicol A, Sorwar E, Villeneuve-Laroche M, Chen ECH, Iwasaki W, Brauer EK, 2023b. Arbuscular mycorrhizal fungi heterokaryons have two nuclear populations with distinct roles in host–plant interactions. Nat Microbiol 8: 2142–2153. 10.1038/s41564-023-01495-8
- ↵St. Pierre CL, Macias-Velasco JF, Wayhart JP, Yin L, Semenkovich CF, Lawson HA. 2022. Genetic, epigenetic, and environmental mechanisms govern allele-specific gene expression. Genome Res 32: 1042–1057. 10.1101/gr.276193.121
- ↵Stukenbrock EH, McDonald BA. 2009. Population genetics of fungal and oomycete effectors involved in gene-for-gene interactions. Mol Plant Microbe Interact 22: 371–380. 10.1094/MPMI-22-4-0371
- ↵Sun H, Jiao W-B, Krause K, Campoy JA, Goel M, Folz-Donahue K, Kukat C, Huettel B, Schneeberger K. 2022. Chromosome-scale and haplotype-resolved genome assembly of a tetraploid potato cultivar. Nat Genet 54: 342–348. 10.1038/s41588-022-01015-0
- ↵Sundararajan K, Straight AF. 2022. Centromere identity and the regulation of chromosome segregation. Front Cell Dev Biol 10: 914249. 10.3389/fcell.2022.914249
- ↵Symonová R. 2019. Integrative rDNAomics—importance of the oldest repetitive fraction of the eukaryote genome. Genes (Basel) 10: 345. 10.3390/genes10050345
- ↵Szymanski M, Zielezinski A, Barciszewski J, Erdmann VA, Karlowski WM. 2016. 5SRNAdb: an information resource for 5S ribosomal RNAs. Nucleic Acids Res 44: D180–D183. 10.1093/nar/gkv1081
- ↵Thach T, Ali S, de Vallavieille-Pope C, Justesen AF, Hovmøller MS. 2016. Worldwide population structure of the wheat rust fungus Puccinia striiformis in the past. Fungal Genet Biol 87: 1–8. 10.1016/j.fgb.2015.12.014
- ↵Tian Y, Thrimawithana A, Ding T, Guo J, Gleave A, Chagné D, Ampomah-Dwamena C, Ireland HS, Schaffer RJ, Luo Z, 2022. Transposon insertions regulate genome-wide allele-specific expression and underpin flower colour variations in apple (Malus spp. Plant Biotechnol J 20: 1285–1297. 10.1111/pbi.13806
- ↵Torres DE, Reckard AT, Klocko AD, Seidl MF. 2023. Nuclear genome organization in fungi: from gene folding to Rabl chromosomes. FEMS Microbiol Rev 47: fuad021. 10.1093/femsre/fuad021
- ↵Upadhyaya NM, Mago R, Panwar V, Hewitt T, Luo M, Chen J, Sperschneider J, Nguyen-Phuc H, Wang A, Ortiz D, 2021. Genomics accelerated isolation of a new stem rust avirulence gene–wheat resistance gene pair. Nat Plants 7: 1220–1228. 10.1038/s41477-021-00971-5
- ↵Varoquaux N, Liachko I, Ay F, Burton JN, Shendure J, Dunham MJ, Vert J-P, Noble WS. 2015. Accurate identification of centromere locations in yeast genomes using Hi-C. Nucleic Acids Res 43: 5331–5339. 10.1093/nar/gkv424
- ↵Vasimuddin M, Misra S, Li H, Aluru S. 2019. Efficient architecture-aware acceleration of BWA-MEM for multicore systems. In 2019 IEEE International Parallel and Distributed Processing Symposium (IPDPS), Rio de Janeiro, Brazil, pp. 314–324. IEEE, Piscataway, NJ. https://ieeexplore.ieee.org/document/8820962 (accessed November 25, 2024). 10.1109/IPDPS.2019.00041
- ↵Vasquez-Gross H, Kaur S, Epstein L, Dubcovsky J. 2020. A haplotype-phased genome of wheat stripe rust pathogen Puccinia striiformis f. sp. tritici, race PST-130 from the western USA. PLoS One 15: e0238611. 10.1371/journal.pone.0238611
- ↵Wallen RM, Perlin MH. 2018. An overview of the function and maintenance of sexual reproduction in dikaryotic fungi. Front Microbiol 9: 503. 10.3389/fmicb.2018.00503
- ↵Wang J, Zhan G, Tian Y, Zhang Y, Xu Y, Kang Z, Zhao J. 2022. Role of sexual reproduction in the evolution of the wheat stripe rust fungus races in China. Phytopathology 112: 1063–1071. 10.1094/PHYTO-08-21-0331-R
- ↵Wang W, Zhang X, Garcia S, Leitch AR, Kovařík A. 2023. Intragenomic rDNA variation - the product of concerted evolution, mutation, or something in between? Heredity (Edinb) 131: 179–188. 10.1038/s41437-023-00634-5
- ↵Wang J, Xu Y, Peng Y, Wang Y, Kang Z, Zhao J. 2024. A fully haplotype-resolved and nearly gap-free genome assembly of wheat stripe rust fungus. Sci Data 11: 508. 10.1038/s41597-024-03361-6
- ↵Wellings CR. 2007. Puccinia striiformis in Australia: a review of the incursion, evolution, and adaptation of stripe rust in the period 1979–2006. Aust J Agric Res 58: 567–575. 10.1071/AR07130
- ↵Xia C, Huang L, Huang J, Zhang H, Huang Y, Benhamed M, Wang M, Chen X, Zhang M, Liu T, 2022. Folding features and dynamics of 3D genome architecture in plant fungal pathogens. Microbiol Spectr 10: e02608-22. 10.1128/spectrum.02608-22
- ↵Xu B, Zeng X-M, Gao X-F, Jin D-P, Zhang L-B. 2017. ITS non-concerted evolution and rampant hybridization in the legume genus Lespedeza (Fabaceae). Sci Rep 7: 40057. 10.1038/srep40057
- ↵Yadav V, Sreekumar L, Guin K, Sanyal K. 2018a. Five pillars of centromeric chromatin in fungal pathogens. PLoS Pathog 14: e1007150. 10.1371/journal.ppat.1007150
- ↵Yadav V, Sun S, Billmyre RB, Thimmappa BC, Shea T, Lintner R, Bakkeren G, Cuomo CA, Heitman J, Sanyal K. 2018b. RNAi is a critical determinant of centromere evolution in closely related fungi. Proc Natl Acad Sci 115: 3108–3113. 10.1073/pnas.1713725115
- ↵Zhao J, Duan W, Xu Y, Zhang C, Wang L, Wang J, Tian S, Pei G, Zhan G, Zhuang H, 2021. Distinct transcriptomic reprogramming in the wheat stripe rust fungus during the initial infection of wheat and barberry. Mol Plant-Microbe Interact MPMI 34: 198–209. 10.1094/MPMI-08-20-0244-R
- ↵Zheng W, Huang L, Huang J, Wang X, Chen X, Zhao J, Guo J, Zhuang H, Qiu C, Liu J, 2013. High genome heterozygosity and endemic genetic recombination in the wheat stripe rust fungus. Nat Commun 4: 2673. 10.1038/ncomms3673