
Haplotype-aware sequence alignment to pangenome graphs
Abstract
Modern pangenome graphs are built using haplotype-resolved genome assemblies. When mapping reads to a pangenome graph, prioritizing
alignments that are consistent with the known haplotypes improves genotyping accuracy. However, the existing rigorous formulations
for colinear chaining and alignment problems do not consider the haplotype paths in a pangenome graph. This often leads to
spurious read alignments to those paths that are unlikely recombinations of the known haplotypes. In this paper, we develop
novel formulations and algorithms for sequence-to-graph alignment and chaining problems. Inspired by the genotype imputation
models, we assume that a query sequence is an imperfect mosaic of reference haplotypes. Accordingly, we introduce a recombination
penalty in the scoring functions for each haplotype switch. First, we solve haplotype-aware sequence-to-graph alignment in
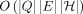 time, where Q is the query sequence, E is the set of edges, and H is the set of haplotypes represented in the graph. To complement our solution, we prove that an
algorithm significantly faster than
time, where Q is the query sequence, E is the set of edges, and H is the set of haplotypes represented in the graph. To complement our solution, we prove that an
algorithm significantly faster than 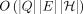 is impossible under the strong exponential time hypothesis (SETH). Second, we propose a haplotype-aware chaining algorithm
that runs in
is impossible under the strong exponential time hypothesis (SETH). Second, we propose a haplotype-aware chaining algorithm
that runs in 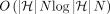 time after graph preprocessing, where N is the count of input anchors. We then establish that a chaining algorithm significantly faster than
time after graph preprocessing, where N is the count of input anchors. We then establish that a chaining algorithm significantly faster than 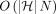 is impossible under SETH. As a proof-of-concept, we implemented our chaining algorithm in the Minichain aligner. By aligning
sequences sampled from the human major histocompatibility complex (MHC) to a pangenome graph of 60 MHC haplotypes, we demonstrate
that our algorithm achieves better consistency with ground-truth recombinations compared with a haplotype-agnostic algorithm.
is impossible under SETH. As a proof-of-concept, we implemented our chaining algorithm in the Minichain aligner. By aligning
sequences sampled from the human major histocompatibility complex (MHC) to a pangenome graph of 60 MHC haplotypes, we demonstrate
that our algorithm achieves better consistency with ground-truth recombinations compared with a haplotype-agnostic algorithm.
Pangenome graphs represent the maximum possible diversity that exists in the population (Singh et al. 2022). A variety of methods have been developed that use pangenome graphs for common applications, including genotyping, variant calling, haplotype reconstruction, metagenomic read classification, etc. (Eizenga et al. 2020). Efforts toward using pangenome graphs have been further catalyzed by the advent of long and accurate reads. The recent progress in long-read technologies and assembly algorithms enables high-quality haplotype-phased assembly of human genomes (Wenger et al. 2019; Cheng et al. 2021; Nurk et al. 2022; Rautiainen et al. 2023). Building a pangenome graph directly from phased assemblies instead of variant calls (VCF files) allows a more comprehensive representation of the variation (Wang et al. 2022). Accordingly, the latest methods for pangenome graph construction use phased assemblies to construct a graph (Li et al. 2020, 2024; Ebler et al. 2022; Garrison et al. 2023; Liao et al. 2023; Hickey et al. 2024). The input haplotypes are stored as paths in the graph.
Pangenome graphs raise fundamental questions about the trade-offs between complexity and usability. An arbitrary path in a pangenome graph corresponds to either the original haplotypes or their recombinations. The number of sequences spelled by a graph increases combinatorially with the number of variants. This issue has been addressed previously by using different techniques, for example, by limiting the amount of variation in the graph (Pritt et al. 2018; Jain et al. 2021; Tavakoli et al. 2022), artificially simplifying complex regions (Garrison et al. 2018), or restricting the alignment to either one (Mokveld et al. 2020; Sirén et al. 2020, 2021) or two haplotype paths (Avila Cartes et al. 2024). A more effective way to address this problem could be to utilize linkage disequilibrium, that is, the correlations between two or more genetic variants (Ebler et al. 2022; Grytten et al. 2022). For example, PanGenie is an alignment-free genotyping algorithm that leverages long-range haplotype information inherent in the phased genomes (Ebler et al. 2022).
A pangenome graph is represented as either a directed cyclic graph or a directed acyclic graph (DAG), where each vertex is
labeled by a sequence. The primary formulation for sequence-to-graph alignment problem seeks a path in the graph that spells
a sequence with minimum edit distance from the query sequence (Manber and Wu 1992; Amir et al. 2000; Navarro 2000; Lee et al. 2002; Rautiainen et al. 2019; Jain et al. 2020; Equi et al. 2023b). O(|V| + |Q||E|)-time algorithms exist for both exact and approximate pattern-matching problems for graphs, where Q denotes the query sequence, E denotes the set of edges, and V denotes the set of vertices. These formulations do not consider the associations between genetic variants and may lead to
alignments with spurious recombinations in variant-dense regions (Pritt et al. 2018). Colinear chaining is another common technique used in modern aligners. It is used to identify a coherent subset of anchors
(short exact matches) that can be joined together to produce an alignment. The existing formulations for chaining on graphs
share the same limitation of not considering the associations between genetic variants (Mäkinen et al. 2019; Chandra and Jain 2023; Ma et al. 2023; Rizzo et al. 2023a; Rajput et al. 2024). Some of these chaining algorithms run in 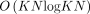 time after graph preprocessing, where K denotes the minimum number of paths required to cover all the vertices, and N denotes the count of input anchors (Chandra and Jain 2023; Ma et al. 2023).
time after graph preprocessing, where K denotes the minimum number of paths required to cover all the vertices, and N denotes the count of input anchors (Chandra and Jain 2023; Ma et al. 2023).
In this paper, we address the above limitations by introducing “haplotype-aware” formulations for (1) sequence-to-DAG alignment
and (2) sequence-to-DAG chaining problems. Our formulations use the haplotype path information available in modern pangenome
graphs. The formulations are inspired from the classic Li–Stephens haplotype copying model (Li and Stephens 2003). The Li–Stephens model is a probabilistic generative model that assumes that a sampled haplotype is an imperfect mosaic
of known haplotypes. Similarly, our formulation for haplotype-aware sequence-to-DAG alignment problem optimizes the number
of edits and haplotype switches simultaneously (Fig. 1). We give the 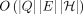 time algorithm for solving the problem, where H denotes the set of haplotypes represented in the pangenome DAG. We further
prove that the existence of a significantly faster algorithm than
time algorithm for solving the problem, where H denotes the set of haplotypes represented in the pangenome DAG. We further
prove that the existence of a significantly faster algorithm than 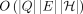 is not possible under the strong exponential time hypothesis (SETH) (Vassilevska Williams 2015). We also formulate the haplotype-aware colinear chaining problem. We solve it in
is not possible under the strong exponential time hypothesis (SETH) (Vassilevska Williams 2015). We also formulate the haplotype-aware colinear chaining problem. We solve it in 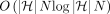 time, assuming that a one-time preprocessing of the DAG is done in
time, assuming that a one-time preprocessing of the DAG is done in 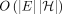 time. To provide evidence of the near optimality of this algorithm, we show that it is impossible to solve this problem significantly
faster than
time. To provide evidence of the near optimality of this algorithm, we show that it is impossible to solve this problem significantly
faster than 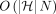 under SETH.
under SETH.
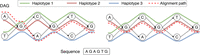
An example of an acyclic pangenome graph built using three haplotypes. The left figure shows the optimal alignment of the query sequence to the DAG with minimum edit distance. Accordingly, the edit distance is zero, and the count of recombinations is two. Next, suppose that we use recombination penalty −5 (formally defined in Methods). The right figure shows the new optimal alignment, where there is no recombination because the alignment path is consistent with haplotype 2.
We implemented the proposed chaining algorithm in Minichain and evaluated it using simulated and real sequences. We built a pangenome DAG using 60 publicly available complete major histocompatibility complex (MHC) sequences (https://zenodo.org/records/6617246). We simulated query MHC haplotypes as mosaics of the 60 haplotypes. We demonstrate that introducing recombination penalty in the formulation leads to a much better consistency between the observed recombination events and the ground truth. We achieved Pearson's correlation up to 0.94 between the observed recombination count and the ground-truth recombination count, whereas the correlation remained below 0.23 if recombinations are not penalized.
We also tested Minichain using publicly available Pacific Biosciences (PacBio) and Oxford Nanopore Technologies (ONT) long reads from the CHM13 human genome (Vollger et al. 2020; Nurk et al. 2022). We demonstrate a better consistency between the ground-truth haplotype (CHM13) and the selected haplotype in the output. Our experiments suggest that haplotype-aware pattern-matching to pangenome graphs can be useful to improve read mapping and variant calling accuracy.
Methods
Haplotype-aware pangenome graphs and sequence alignments
First, we present the background concepts and notations before introducing our problem formulations and algorithms.
Haplotype-aware pangenome graph
Let 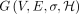 be a DAG representing a pangenome reference. We assume that the DAG is weakly connected and |E| ≥ |V| − 1. Let Σ denote an alphabet. Function
be a DAG representing a pangenome reference. We assume that the DAG is weakly connected and |E| ≥ |V| − 1. Let Σ denote an alphabet. Function 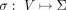 assigns a character label to each vertex.
assigns a character label to each vertex. 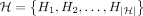 denotes the set of haplotype sequences represented in the DAG. Each haplotype sequence can be identified using a path in
the DAG. We define haps(v) as {i|Hi includes vertex v}. We assume that haps(v) ≠ ϕ for all v ∈ V and
denotes the set of haplotype sequences represented in the DAG. Each haplotype sequence can be identified using a path in
the DAG. We define haps(v) as {i|Hi includes vertex v}. We assume that haps(v) ≠ ϕ for all v ∈ V and 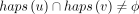 for all edges (u, v) ∈ E. In other words, each vertex and edge must be supported by at least one haplotype. Given a query sequence Q ∈ Σ+, we will find its exact and approximate matches in the DAG. For brevity, we use [m] to denote the set {1, 2, …, m}, m ∈ ℕ. Let an alignment path of length l in the DAG be denoted as an ordered sequence (w1, w2, …, wl), where each wi is a pair of the form (v, h) such that v ∈ V, h ∈ haps(v), and (wi.v, wi+1.v) ∈ E for all i ∈ [l − 1]. Thus, in our haplotype-aware setting, an alignment path specifies a path in the DAG and the indices of the selected
haplotypes along the path. We say that a recombination has occurred in between wi and wi+1 if wi.h ≠ wi+1.h. Given an alignment path P in the DAG, we use functions
for all edges (u, v) ∈ E. In other words, each vertex and edge must be supported by at least one haplotype. Given a query sequence Q ∈ Σ+, we will find its exact and approximate matches in the DAG. For brevity, we use [m] to denote the set {1, 2, …, m}, m ∈ ℕ. Let an alignment path of length l in the DAG be denoted as an ordered sequence (w1, w2, …, wl), where each wi is a pair of the form (v, h) such that v ∈ V, h ∈ haps(v), and (wi.v, wi+1.v) ∈ E for all i ∈ [l − 1]. Thus, in our haplotype-aware setting, an alignment path specifies a path in the DAG and the indices of the selected
haplotypes along the path. We say that a recombination has occurred in between wi and wi+1 if wi.h ≠ wi+1.h. Given an alignment path P in the DAG, we use functions 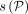 and
and 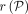 to denote the sequence spelled by the path and the count of recombinations in the path, respectively.
to denote the sequence spelled by the path and the count of recombinations in the path, respectively.
Hardness result for the edit distance problem
Our hardness result (Lemma 3) will build on top of the seminal work of Backurs and Indyk (2015) . The orthogonal vector (OV) problem asks the following: given two sets of d-dimensional binary vectors A and B, |A| = |B| =: n, decide if there is a pair a ∈ A, b ∈ B such that a · b = 0. If the SETH (Vassilevska Williams 2015) holds and 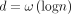 , there is no
, there is no 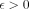 for which an
for which an 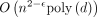 algorithm can solve the OV problem (Williams 2005). This result is frequently used to study the hardness of other problems (Backurs and Indyk 2015; Hoppenworth et al. 2020; Gibney et al. 2022; Equi et al. 2023a,b). For any two sequences P1 and P2, let EDIT(P1, P2) be the edit distance between them. Let us define another distance measure between two sequences as following:
algorithm can solve the OV problem (Williams 2005). This result is frequently used to study the hardness of other problems (Backurs and Indyk 2015; Hoppenworth et al. 2020; Gibney et al. 2022; Equi et al. 2023a,b). For any two sequences P1 and P2, let EDIT(P1, P2) be the edit distance between them. Let us define another distance measure between two sequences as following: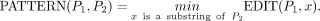 Backurs and Indyk (2015) proved the hardness of computing edit distance by designing two reductions: (1) OV to PATTERN and (2) PATTERN to EDIT. We
recall the properties of their first reduction gadget.
Backurs and Indyk (2015) proved the hardness of computing edit distance by designing two reductions: (1) OV to PATTERN and (2) PATTERN to EDIT. We
recall the properties of their first reduction gadget.
(Backurs and Indyk 2015): Given an OV problem instance with sets A = {a1, …, an} and B = {b1, …, bn} comprising d-dimensional binary vectors, there exist transformations of sets A and B into two sequences S1 and S2, respectively, over the alphabet {0, 1, 2} such that the following properties hold:
-
– Computing PATTERN(S1, S2) determines the existence of orthogonal vectors. There exist constants β1, β2 ∈ ℕ such that if there is a pair of orthogonal vectors a ∈ A, b ∈ B, then PATTERN(S1, S2) ≤ β1n − β2; otherwise, PATTERN(S1, S2) = β1n.
-
– |S1| is a function of n and d. Similarly, |S2| is a function of n and d. Both |S1| and |S2| are in O(n · poly(d)).
-
– Time to compute sequences S1 and S2 from sets A and B is in O(n · poly(d)).
We denote the injective function that generates the gadget sequence S1 from set A as f1. Similarly, injective function f2 generates sequence S2 from set B. The exact definitions of f1 and f2 are available elsewhere (Backurs and Indyk 2015).
Problem formulations for haplotype-aware sequence alignment
The input to each of the following problem is a DAG 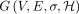 , a query Q ∈ Σ+, and a user-specified threshold k ∈ ℕ. In each problem, we seek a match of the query in the DAG while controlling the number of recombinations.
, a query Q ∈ Σ+, and a user-specified threshold k ∈ ℕ. In each problem, we seek a match of the query in the DAG while controlling the number of recombinations.
-
– Problem 1 (exact matching, limited recombinations): Determine if there is an alignment path P in DAG G such that
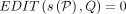 and
and 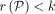 .
.
-
– Problem 2 (approximate matching, no recombination): Determine if there is an alignment path P in DAG G such that
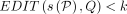 and
and 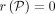 .
.
-
– Problem 3 (approximate matching, limited recombinations): Let k, c1, c2 be user-specified thresholds, where k ∈ ℕ and c1, c2 ∈ ℝ ≥ 0. Here c1 indicates cost per edit, and c2 indicates cost per recombination. Determine if there is an alignment path P in DAG G such that
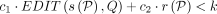 .
.
In Problem 3, we assume a constant cost per recombination but one can also consider using different cost per locus based on known rates of recombination (Petes 2001). In the above set of problems, we have omitted the case with exact matching and no recombination. One way to solve this case is by doing exact sequence matching against each haplotype independently.
Proposed algorithms and theoretical analysis
Problems 1–3 can be solved in 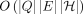 time.
time.
We show how to solve Problem 3 using dynamic programming. The same can be extended to Problems 1 and 2 as well. Our approach
is similar to the standard sequence-to-DAG alignment algorithm (Lee et al. 2002) except that we also track recombinations. We add a dummy source vertex,  , with an empty label. It has zero incoming edges and |V| outgoing edges to all v ∈ V . Assume
, with an empty label. It has zero incoming edges and |V| outgoing edges to all v ∈ V . Assume 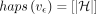 . Let D(i, v, h) denote the minimum cost for aligning the (possibly empty) prefix of sequence Q ending at index i against an alignment path that ends at pair (v, h),
. Let D(i, v, h) denote the minimum cost for aligning the (possibly empty) prefix of sequence Q ending at index i against an alignment path that ends at pair (v, h), 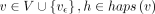 . We also maintain a second table,
. We also maintain a second table, 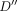 , such that
, such that 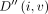 will store
will store 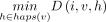 . Initialize
. Initialize 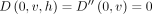 for all
for all 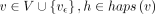 . Initialize the remaining cells in table
. Initialize the remaining cells in table 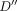 to ∞. For
to ∞. For 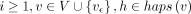 , update D(i, v, h) and
, update D(i, v, h) and 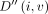 using the following recursion:
using the following recursion: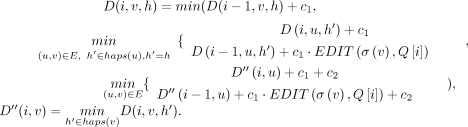 The recursion computes the optimal value of D(i, v, h) by considering the possibility of character deletion, insertion, match, and mismatch. The product c1 · EDIT(σ(v), Q[i]) equals zero if there is a match between characters σ(v) and Q[i]. It equals c1 in case of a mismatch. The other expressions involve addition of c1 to consider the possibility of an insertion or a deletion. The expressions involving the addition of c2 evaluate the possibility of a recombination. After updating D(i, v, h) for all h ∈ haps(v), we update
The recursion computes the optimal value of D(i, v, h) by considering the possibility of character deletion, insertion, match, and mismatch. The product c1 · EDIT(σ(v), Q[i]) equals zero if there is a match between characters σ(v) and Q[i]. It equals c1 in case of a mismatch. The other expressions involve addition of c1 to consider the possibility of an insertion or a deletion. The expressions involving the addition of c2 evaluate the possibility of a recombination. After updating D(i, v, h) for all h ∈ haps(v), we update 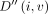 using the above equation. We compute values in table D in the increasing order of i and then the increasing topological order of
using the above equation. We compute values in table D in the increasing order of i and then the increasing topological order of 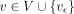 for a fixed i. The output, namely, the minimum alignment cost, is
for a fixed i. The output, namely, the minimum alignment cost, is 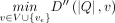 . The runtime of the algorithm is dominated by the recursion. Solving the recursion takes
. The runtime of the algorithm is dominated by the recursion. Solving the recursion takes 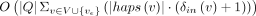 time, where δin(v) denotes the in-degree of vertex v. As a result, the time complexity of the algorithm is
time, where δin(v) denotes the in-degree of vertex v. As a result, the time complexity of the algorithm is 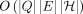 .
.
Next, we prove that a significantly faster algorithm than 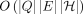 for solving Problem 2 or 3 is unlikely. The existence of a faster algorithm for Problem 1 remains open.
for solving Problem 2 or 3 is unlikely. The existence of a faster algorithm for Problem 1 remains open.
For any constant 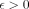 , there is no algorithm that solves Problem 2 or Problem 3 in
, there is no algorithm that solves Problem 2 or Problem 3 in 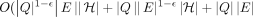
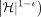 time, unless SETH is false.
time, unless SETH is false.
Problem 3 is a generalized version of Problem 2. Therefore, it suffices to prove the hardness of Problem 2. We derive the
reduction from an OV problem. Suppose we have an OV instance with equal-sized sets A and B comprising d-dimensional binary vectors. Define n = |A| = |B|. We assume w.l.o.g. that n is a perfect square. We partition set A into 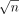 subsets
subsets  , each containing
, each containing 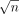 vectors. Similarly, we partition set B into
vectors. Similarly, we partition set B into 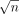 subsets
subsets 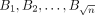 , each containing
, each containing 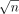 vectors. Observe that solving the OV problem for all (Ai, Bj) instances,
vectors. Observe that solving the OV problem for all (Ai, Bj) instances, 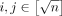 , solves the OV problem for instance (A, B).
, solves the OV problem for instance (A, B).
Next, we describe the construction of our gadget comprising query sequences 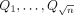 and haplotype-aware pangenome DAG
and haplotype-aware pangenome DAG 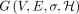 . Assume the alphabet {0, 1, 2} for defining the query sequences and the vertex labels in the DAG. Define Qi = f1(Ai) for all
. Assume the alphabet {0, 1, 2} for defining the query sequences and the vertex labels in the DAG. Define Qi = f1(Ai) for all 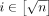 (ref. Lemma 1). Next, we will construct
(ref. Lemma 1). Next, we will construct 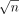 haplotype sequences. Define Hi = f2(Bi) for all
haplotype sequences. Define Hi = f2(Bi) for all 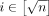 . Using Lemma 1, we know that all our haplotype sequences have uniform length; that is,
. Using Lemma 1, we know that all our haplotype sequences have uniform length; that is, 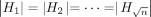 . Next, we construct a DAG where these haplotype sequences can be represented. We build a DAG with |Hi| “layers.” Each layer comprises three vertices labeled with characters “0,” “1,” and “2,” respectively (see Fig. 2A). Each layer (except the last) is fully connected to its next adjacent layer using 32 = 9 directed edges. Subsequently, we identify the unique path that spells each of the
. Next, we construct a DAG where these haplotype sequences can be represented. We build a DAG with |Hi| “layers.” Each layer comprises three vertices labeled with characters “0,” “1,” and “2,” respectively (see Fig. 2A). Each layer (except the last) is fully connected to its next adjacent layer using 32 = 9 directed edges. Subsequently, we identify the unique path that spells each of the 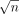 haplotype sequences in this DAG. Finally, we discard the vertices and edges that are not used by any haplotype sequence (Fig. 2B).
haplotype sequences in this DAG. Finally, we discard the vertices and edges that are not used by any haplotype sequence (Fig. 2B).
Next, the above gadget will allow us to solve the OV problem by invoking the haplotype-aware sequence to graph alignment algorithm
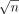 times, once for each of the
times, once for each of the  query sequences. An alignment path with no recombination in the DAG spells a substring of a haplotype. Using Lemma 1, we
make the following claim. There exist the two orthogonal vectors a ∈ A and b ∈ B if and only if we have at least one query sequence
query sequences. An alignment path with no recombination in the DAG spells a substring of a haplotype. Using Lemma 1, we
make the following claim. There exist the two orthogonal vectors a ∈ A and b ∈ B if and only if we have at least one query sequence 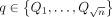 for which an alignment path P exists such that
for which an alignment path P exists such that 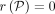 and
and 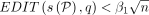 . It is only remaining to show that a faster algorithm for Problem 2 will contradict SETH by enabling a faster algorithm for
the OV problem. Observe that (1) each |Qi| and |Hi| is in
. It is only remaining to show that a faster algorithm for Problem 2 will contradict SETH by enabling a faster algorithm for
the OV problem. Observe that (1) each |Qi| and |Hi| is in 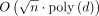 ; (2) |V| and |E| are asymptotically equal to |Hi|; (3) number of haplotypes is
; (2) |V| and |E| are asymptotically equal to |Hi|; (3) number of haplotypes is 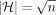 ; and (4) we invoke the alignment algorithm
; and (4) we invoke the alignment algorithm 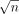 times. Construction of the gadget requires O(n · poly(d)) time. Therefore, if Problem 2 can be solved in
times. Construction of the gadget requires O(n · poly(d)) time. Therefore, if Problem 2 can be solved in 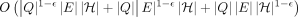 time for some
time for some 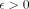 , then the OV problem can be solved in
, then the OV problem can be solved in 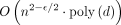 time.
time.
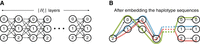
An illustration of the pangenome DAG used in the reduction proof of Lemma 3. An initial version of the DAG (A) and the changes made after addition of all the haplotype paths (B).
Methods for haplotype-aware chaining on graphs
Most alignment tools use a seed-chain-extend heuristic to compute the alignments quickly (Sahlin et al. 2023; Shaw and Yu 2023). Given a set of seed matches (anchors) as input, colinear chaining is a rigorous optimization technique to identify promising alignment regions in a reference. It identifies a coherent subset of anchors such that their coordinates are ordered on the query and the reference. The selected anchors are subsequently combined together to form an alignment. Several versions of colinear chaining problems have been studied for aligning two sequences (Eppstein et al. 1992a,b; Myers and Miller 1995; Abouelhoda and Ohlebusch 2005; Otto et al. 2011; Mäkinen and Sahlin 2020; Jain et al. 2022). Recent works have further studied the extension of chaining on acyclic (Mäkinen et al. 2019; Chandra and Jain 2023; Ma et al. 2023; Rizzo et al. 2023a) and cyclic pangenome graphs (Antipov et al. 2016; Rajput et al. 2024), but these formulations do not consider the haplotype paths.
Concepts and notations for haplotype-aware colinear chaining
From here on, we assume that vertices of a haplotype-aware pangenome DAG are labeled with sequences. Sequence-labeled vertices
permit a more compact graph representation, which will be useful in the context of chaining. It is trivial to transform a
graph from sequence-labeled form to character-labeled form and vice versa. Let function σ:V → Σ+ label each vertex v ∈ V with sequence σ(v) in DAG 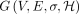 . The input to the chaining problem is a set of anchors, {M[1], M[2], …, M[N]}. An anchor is represented by a three-tuple (v, [x..y], [c..d]), which signifies a match between query substring Q[c..d] and substring σ(v)[x..y] in the DAG (Fig. 3). The weight function assigns a user-specified weight to each anchor. Anchor weights are typically set proportional to the length of the
matching substrings in practice. We use R−(v)⊆ V to denote the subset of vertices that can reach v using a path in the DAG. Vertex v is also included in R−(v). Next, we define the precedence relationship between a pair of anchors.
. The input to the chaining problem is a set of anchors, {M[1], M[2], …, M[N]}. An anchor is represented by a three-tuple (v, [x..y], [c..d]), which signifies a match between query substring Q[c..d] and substring σ(v)[x..y] in the DAG (Fig. 3). The weight function assigns a user-specified weight to each anchor. Anchor weights are typically set proportional to the length of the
matching substrings in practice. We use R−(v)⊆ V to denote the subset of vertices that can reach v using a path in the DAG. Vertex v is also included in R−(v). Next, we define the precedence relationship between a pair of anchors.
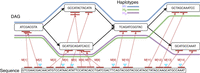
An example that shows anchors (in red) between a query sequence and a pangenome DAG. The DAG comprises three haplotype sequences, H1, H2, H3. In this example, the ordered sequence of pairs [(2, 1), (4, 1), (5, 1), (9, 1), (10, 1), (13, 3), (14, 3)] forms a chain, and the corresponding anchors are highlighted using blue markers. This chain includes a single recombination.
Given two anchors M[i] and M[j], M[i] precedes (≺) M[j] if (1) M[i].d < M[j].c, (2) M[i].v ∈ R−(M[j].v), and (3) M[i].y < M[j].x if M[i].v = M[j].v.
A chain is an ordered sequence (s1, s2, …, sp), where each sj is a pair of the form (i, h) such that i ∈ [N], h ∈ haps(M[i].v), and 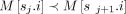 for all j ∈ [p − 1].
for all j ∈ [p − 1].
In our definition, a chain specifies the indices of the selected haplotypes alongside the anchors (Fig. 3). For the sake of efficiency, our precedence condition does not permit overlap between adjacent anchors in a chain. We say that a recombination has occurred in between sj and sj+1 if sj.h ≠ sj+1.h. Given a chain S, we use the function r(S) to denote the count of recombinations in S. Let γ ∈ ℝ≥0 be a user-specified parameter that specifies cost per recombination.
Similar to work by Mäkinen et al. (2019), we will use a path cover to facilitate efficient sparse dynamic programming on the DAG. A path cover is a set of paths in the DAG such that every
vertex in V is included in at least one path. Previous chaining algorithms (see, e.g., Mäkinen et al. 2019; Chandra and Jain 2023; Ma et al. 2023) identify a path cover with the minimum number of paths because the runtime of sparse dynamic programming increases with
the size of the path cover (Mäkinen et al. 2019). In the haplotype-aware setting, we will directly use the paths corresponding to the haplotype sequences as path cover.
Let  be the paths corresponding to the haplotype sequences
be the paths corresponding to the haplotype sequences  , respectively.
, respectively.  is a path cover of the DAG because each vertex in the DAG is included in at least one haplotype (Methods subsection “Haplotype-aware
pangenome graph”). Suppose
is a path cover of the DAG because each vertex in the DAG is included in at least one haplotype (Methods subsection “Haplotype-aware
pangenome graph”). Suppose 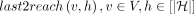 denotes the last vertex in path Ph that belongs to R−(v). last2reach(v, h) does not exist if no vertex in path Ph can reach vertex v. We precompute last2reach(v, h) for all v ∈ V and
denotes the last vertex in path Ph that belongs to R−(v). last2reach(v, h) does not exist if no vertex in path Ph can reach vertex v. We precompute last2reach(v, h) for all v ∈ V and 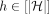 in
in 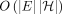 time by using the algorithm from Mäkinen et al. (2019). This is a one-time preprocessing step that will be useful to solve the chaining problems efficiently. Next, we also use
the following standard data structure in our chaining algorithm.
time by using the algorithm from Mäkinen et al. (2019). This is a one-time preprocessing step that will be useful to solve the chaining problems efficiently. Next, we also use
the following standard data structure in our chaining algorithm.
(Mäkinen et al. 2015): Let n be the number of leaves in a tree, each storing a (key, value) pair. The data structure uses a balanced binary search tree. It supports update and range maximum query (RMQ) operations, defined below, in 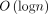 time:
time:
-
– update(k, val): For the leaf w with key = k,
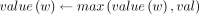 .
.
-
– RMQ(l, r): Return
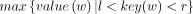 .
.
Given n (key, value) pairs, the tree can be constructed in 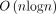 time and O(n) space.
time and O(n) space.
Problem formulations for haplotype-aware colinear chaining
-
– Problem 4 (limited recombinations): Determine an optimal chain S = s1, s2, …, sp that maximizes the chaining score, defined as
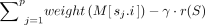 .
.
The second term γ · r(S) in the above scoring function corresponds to the penalty for haplotype switches in a chain. While scoring a chain, it is also essential to penalize the gap corresponding to the distance (in base pairs) between adjacent pair of anchors. Accordingly, we formulate a gap-sensitive version of the above problem. We assume the same definition of function gap(M[i], M[j]), i, j ∈ [N] as used by Chandra and Jain (2023); see Problem 2a in their paper.
-
– Problem 5 (limited recombinations, gap-sensitive): Determine an optimal chain S = s1, s2, …, sp that maximizes the chaining score, defined as
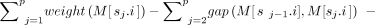
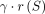 .
.
Proposed chaining algorithms and theoretical analysis
First, it is useful to discuss a simple dynamic programming solution for Problem 4. Let C(i, h) denote the optimal score of a chain ending at pair (i, h), where i ∈ [N], h ∈ haps(M[i].v). We use 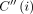 to store maxh∈haps(M[i].v)C(i, h). A single anchor is a valid chain of size one; therefore, we initialize C(i, h) to weight(M[i]) for all i ∈ [N], h ∈ haps(M[i].v). Also, initialize
to store maxh∈haps(M[i].v)C(i, h). A single anchor is a valid chain of size one; therefore, we initialize C(i, h) to weight(M[i]) for all i ∈ [N], h ∈ haps(M[i].v). Also, initialize 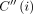 to weight(M[i]) for all i ∈ [N]. We update C(i, h) to its optimal value using the following recursion:
to weight(M[i]) for all i ∈ [N]. We update C(i, h) to its optimal value using the following recursion: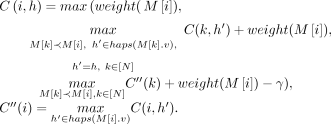 If
If 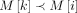 and path Ph covers both the vertices M[k].v and M[i].v, then the chaining score obtained using C(k, h) is considered without recombination penalty. The third term in the expression considers the score with recombination penalty
γ. This is needed when
and path Ph covers both the vertices M[k].v and M[i].v, then the chaining score obtained using C(k, h) is considered without recombination penalty. The third term in the expression considers the score with recombination penalty
γ. This is needed when 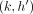 ,
, 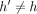 precedes (i, h). After updating C(i, h) for all h ∈ haps(M[i].v), we also update
precedes (i, h). After updating C(i, h) for all h ∈ haps(M[i].v), we also update 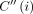 using the above equation. Let function rank(v) specify the topological ordering of vertex v in the DAG. The C(i, h) values should be computed in lexicographically ascending order based on the key (rank(M[i].v), M[i].x). For a fixed i, C(i, h) values may be computed in any order. The above algorithm fills up to
using the above equation. Let function rank(v) specify the topological ordering of vertex v in the DAG. The C(i, h) values should be computed in lexicographically ascending order based on the key (rank(M[i].v), M[i].x). For a fixed i, C(i, h) values may be computed in any order. The above algorithm fills up to 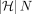 values in table C. Computing each value using a linear scan would require considering all N number of anchors and filtering those that satisfy the precedence criteria. Even if we ignore the time for filtering and
checking precedence, the worst-case asymptotic runtime of this algorithm grows at least as fast as
values in table C. Computing each value using a linear scan would require considering all N number of anchors and filtering those that satisfy the precedence criteria. Even if we ignore the time for filtering and
checking precedence, the worst-case asymptotic runtime of this algorithm grows at least as fast as 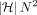 . This makes it too slow when N is in millions, which typically happens when aligning megabase-long de novo assembled sequences to human pangenome DAGs.
To address this, we propose a faster
. This makes it too slow when N is in millions, which typically happens when aligning megabase-long de novo assembled sequences to human pangenome DAGs.
To address this, we propose a faster 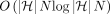 time algorithm for solving Problem 4. First, we note the following inequality.
time algorithm for solving Problem 4. First, we note the following inequality.
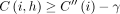 for all i ∈ [N], h ∈ haps(M[i].v).
for all i ∈ [N], h ∈ haps(M[i].v).
Consider any value of i ∈ [N] and h ∈ haps(M[i].v). Let 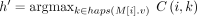 , breaking ties arbitrarily if necessary. Then,
, breaking ties arbitrarily if necessary. Then, 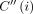 equals
equals 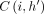 . If
. If 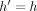 , then
, then 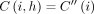 ; thus, the inequality holds. Next, consider the case when
; thus, the inequality holds. Next, consider the case when 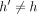 . Let S1 = s1, s2, …, sp−1, sp be an optimal chain that ends at pair
. Let S1 = s1, s2, …, sp−1, sp be an optimal chain that ends at pair 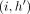 . We consider another chain,
. We consider another chain,  , such that
, such that 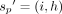 . Suppose the score of chain S2 is x. Both chains S1 and S2 differ only by the haplotype used in their last pairs. Therefore, the difference in the chaining scores of S1 and S2 is at most γ (one recombination). Thus,
. Suppose the score of chain S2 is x. Both chains S1 and S2 differ only by the haplotype used in their last pairs. Therefore, the difference in the chaining scores of S1 and S2 is at most γ (one recombination). Thus, 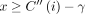 . Also, C(i, h) must be ≥x.
. Also, C(i, h) must be ≥x.
Assuming DAG 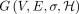 is preprocessed, Problem 4 can be solved in
is preprocessed, Problem 4 can be solved in 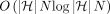 time.
time.
See Algorithm 1 for an outline of our approach. We extend the sparse dynamic programming framework of Mäkinen et al. (2019) to our haplotype-aware setting. Our path cover  contains
contains 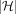 paths corresponding to the
paths corresponding to the 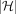 haplotype sequences. We use
haplotype sequences. We use 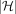 search trees
search trees  , one per path. Each search tree
, one per path. Each search tree 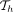 will record C(i, h) scores specific to that haplotype.
will record C(i, h) scores specific to that haplotype.
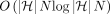 time chaining algorithm for Problem 4
time chaining algorithm for Problem 4
Input: Weighted anchors M[1..N], haplotype paths  , last2reach values, parameter γ
, last2reach values, parameter γ
Output: Table C such that C(i, h)= optimal score of a chain that ends at (i, h), for all 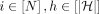
1 Initialize search trees 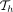 , for all
, for all 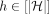 , using keys {M[i].d|1 ≤ i ≤ N} and values −∞
, using keys {M[i].d|1 ≤ i ≤ N} and values −∞
2 Initialize 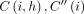 and Cl2r(i) as weight(M[i]), for all
and Cl2r(i) as weight(M[i]), for all 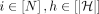
3 /* Create array Z that stores tuples of the form (v, pos, task, anchor, haplotype), where v ∈ V, pos ∈ ℕ, task ∈ {0, 1, 2}, anchor ∈ [N], and 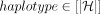 .*/
.*/
4 for i ← 1 to N do
5 for 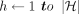 do
do
6 if h ∈ haps(M[i].v) then
7 Z.push(M[i].v, M[i].x, 0, i, h)
8 Z.push(M[i].v, M[i].x, 1, i, h)
9 Z.push(M[i].v, M[i].y, 2, i, h)
10 else if last2reach(M[i].v, h) exists then
11 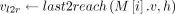
12 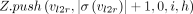
13 end
14 end
15 for z ∈ Z in lexicographically ascending order based on the key (rank(v), pos, task) do
16 i ← z.anchor, h ← z.haplotype, v ← z.v, wt ← weight(M[i])
17 if z.task = 0 then
18 if h ∈ haps(M[i].v) then
19 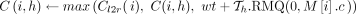
20 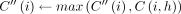
21 else
22 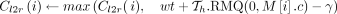
23 else if z.task = 1 then
24 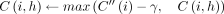
25 else if z.task = 2 then
26 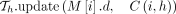
27 end
We initialize each search tree with keys {M[i].d |1 ≤ i ≤ N} and values −∞ (Line 1). In Lines 4–13, we create an array Z with 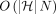 tuples. After sorting of Z (Line 15), these tuples ensure that the search trees are updated and queried in a proper order. At the time of processing
C(i, h), i ∈ [N], h ∈ haps(M[i].v), the algorithm would have already finished processing
tuples. After sorting of Z (Line 15), these tuples ensure that the search trees are updated and queried in a proper order. At the time of processing
C(i, h), i ∈ [N], h ∈ haps(M[i].v), the algorithm would have already finished processing 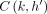 for all
for all 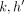 such that
such that 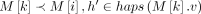 . The score
. The score 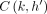 would have already been recorded in search tree
would have already been recorded in search tree 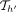 (Line 26).
(Line 26).
All the anchors that lack a preceding anchor are optimally processed at the initialization stage (Line 2). Suppose an optimal
chain ending at (i, h) is s1, s2, …, sp−1, sp such that 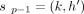 and sp = (i, h). Assume that
and sp = (i, h). Assume that 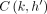 is already computed optimally. While calculating C(i, h), the algorithm handles the following three cases:
is already computed optimally. While calculating C(i, h), the algorithm handles the following three cases:
-
– (Case 1)
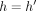 : In this case, we optimally compute C(i, h) by executing a range query in search tree Th (Line 19). Because
: In this case, we optimally compute C(i, h) by executing a range query in search tree Th (Line 19). Because 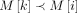 and
and 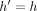 , search tree Th returns the value
, search tree Th returns the value 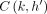 stored in key M[k].d.
stored in key M[k].d.
-
– (Case 2)
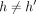 , and path
, and path 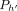 does not cover M[i].v: Observe that
does not cover M[i].v: Observe that 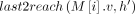 must exist in this case because M[k].v must reach M[i].v. In Lines 10–12, we add tuples in array Z to ensure that Cl2r(i) is updated by using a range query in search tree
must exist in this case because M[k].v must reach M[i].v. In Lines 10–12, we add tuples in array Z to ensure that Cl2r(i) is updated by using a range query in search tree 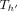 (Line 22). We also put a penalty γ owing to recombination in this case. This update to Cl2r(i) happens after all anchors in vertex
(Line 22). We also put a penalty γ owing to recombination in this case. This update to Cl2r(i) happens after all anchors in vertex 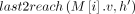 have been processed and search tree
have been processed and search tree 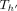 has been updated because of our definition and ordering of the tuples in array Z (Lines 12, 15). Later when we update C(i, h) in Line 19, it gets its optimal value using Cl2r(i).
has been updated because of our definition and ordering of the tuples in array Z (Lines 12, 15). Later when we update C(i, h) in Line 19, it gets its optimal value using Cl2r(i).
-
– (Case 3)
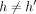 , and path
, and path 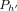 covers M[i].v: If
covers M[i].v: If 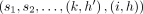 is an optimal chain ending at (i, h), then
is an optimal chain ending at (i, h), then 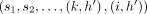 must be an optimal chain that ends at
must be an optimal chain that ends at 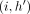 . Accordingly,
. Accordingly, 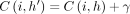 . It implies
. It implies 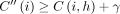 . Using the inequality in Lemma 5, we get
. Using the inequality in Lemma 5, we get 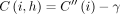 . Accordingly, C(i, h) is updated to its optimal value in Line 24.
. Accordingly, C(i, h) is updated to its optimal value in Line 24.
The total runtime of the algorithm is dominated by sorting of array Z. Sorting an array of size 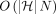 takes
takes 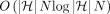 time. Initializing
time. Initializing 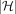 search trees takes
search trees takes 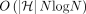 time. The algorithm uses
time. The algorithm uses 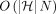 number of
number of 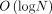 -time RMQ and update operations on the search trees (Lemma 4).
-time RMQ and update operations on the search trees (Lemma 4).
In the above algorithm, the optimal chain can be obtained easily by adding backtracking pointers. We next show that any algorithm
to solve Problem 4 has a time complexity that is not polynomially faster than 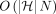 under SETH.
under SETH.
For any constant 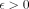 , there is no algorithm that solves Problem 4 in
, there is no algorithm that solves Problem 4 in 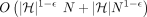 time unless SETH is false.
time unless SETH is false.
The reduction from the OV problem is as follows. Suppose an OV instance is given as A = {a1, …, an} and B = {b1, …, bn} and each set is comprised of d-dimensional binary vectors.
We perform this reduction by constructing a haplotype-aware graph based on A and a sequence based on B with anchors such that there is an anchor chain of some desired value if and only if there is an orthogonal pair of vectors. The main intuition is that an orthogonal pair of vectors should allow a high-value portion of the chain between the corresponding vector gadgets, whereas the remaining vector gadgets can be used in some portion of the chain and give a predictable value.
To this end, we first define component gadgets: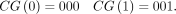 We use
We use 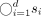 to denote the concatenation of sequences s1, …, sd and
to denote the concatenation of sequences s1, …, sd and 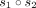 to denote the concatenation of two sequences, s1 and s2. For a symbol c and an integer i, we use ci to denote symbol c repeated i times. Using the above component gadgets, we define our vector gadgets:
to denote the concatenation of two sequences, s1 and s2. For a symbol c and an integer i, we use ci to denote symbol c repeated i times. Using the above component gadgets, we define our vector gadgets: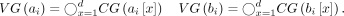 The query sequence is defined as
The query sequence is defined as 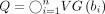 . For 1 ≤ i ≤ n, we construct haplotype sequence Hi from vector ai:
. For 1 ≤ i ≤ n, we construct haplotype sequence Hi from vector ai: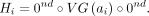 To create the vertices of graph G, we start with the following: a vertex denoted vleft with label 0nd; 2d vertices denoted vx,0, vx,1 for 1 ≤ x ≤ d, where each vx,0 has label CG(0) and each vx,1 has label CG(1); and a vertex denoted vright with label 0nd. For edges, we add edges from vleft to v1,0 and from vleft to v1,1; for 1 ≤ x < d, we add edges from vx,0 to vx+1,0 and vx+1,1 and from vx,1 to vx+1,0 and vx+1,1; and we then add edges from vd,0 to vright and from vd,1 to vright. Next, we embed each haplotype in G by following its only possible path from vleft to vright and deleting any vertices and edges not supported by some haplotype. See Figure 4. We call the subgraph of G excluding only vleft and vright the center of G.
To create the vertices of graph G, we start with the following: a vertex denoted vleft with label 0nd; 2d vertices denoted vx,0, vx,1 for 1 ≤ x ≤ d, where each vx,0 has label CG(0) and each vx,1 has label CG(1); and a vertex denoted vright with label 0nd. For edges, we add edges from vleft to v1,0 and from vleft to v1,1; for 1 ≤ x < d, we add edges from vx,0 to vx+1,0 and vx+1,1 and from vx,1 to vx+1,0 and vx+1,1; and we then add edges from vd,0 to vright and from vd,1 to vright. Next, we embed each haplotype in G by following its only possible path from vleft to vright and deleting any vertices and edges not supported by some haplotype. See Figure 4. We call the subgraph of G excluding only vleft and vright the center of G.
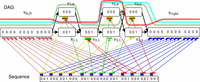
Reduction from OV problem with sets A = {0110, 1100, 1101} and B = {1010, 1001, 1011}.
The anchors are described next. For all i ∈ [n] and for all x ∈ [d], we define the following weight-1 anchors: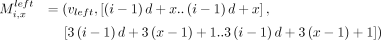 We similarly define weight-1 anchors:
We similarly define weight-1 anchors: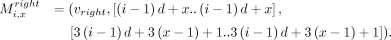 For all i ∈ [n] and for all x ∈ [d], if bi[x] = 0 and if vx,0 and vx,1 exist, we define the following weight-2 anchors
For all i ∈ [n] and for all x ∈ [d], if bi[x] = 0 and if vx,0 and vx,1 exist, we define the following weight-2 anchors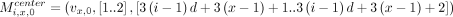
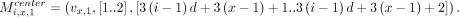 If either vx,0 or vx,1 do not exist, we omit the corresponding anchor. If bi[x] = 1 and vx,0 exists, we define the following weight-2 anchor:
If either vx,0 or vx,1 do not exist, we omit the corresponding anchor. If bi[x] = 1 and vx,0 exists, we define the following weight-2 anchor: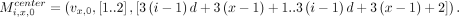 We set the recombination penalty γ = 1. We show the correctness of the above reduction in Lemmas 8–11.
We set the recombination penalty γ = 1. We show the correctness of the above reduction in Lemmas 8–11.
If there exists an orthogonal pair ai ∈ A and bj ∈ B, then there exists an anchor chain with cost 2d + (n − 1)d.
First take anchors 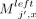 for
for 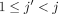 , 1 ≤ x ≤ d. This part of the chain has a value of (j − 1)d. Next, for 1 ≤ x ≤ d, if ai[x] = 0 and bj[x] = 0 or ai[x] = 0 and bj[x] = 1, take
, 1 ≤ x ≤ d. This part of the chain has a value of (j − 1)d. Next, for 1 ≤ x ≤ d, if ai[x] = 0 and bj[x] = 0 or ai[x] = 0 and bj[x] = 1, take 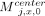 ; if ai[x] = 1 and bj[x] = 0, take
; if ai[x] = 1 and bj[x] = 0, take 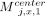 . This part of the chain has a value of 2d. Lastly, for
. This part of the chain has a value of 2d. Lastly, for 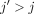 , 1 ≤ x ≤ d, take anchors
, 1 ≤ x ≤ d, take anchors 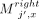 ; this part of the chain has value (n − j)d. This forms a valid chain and requires no recombinations. Thus, the total score is (j − 1)d + 2d + (n − j)d = 2d + (n − 1)d.
; this part of the chain has value (n − j)d. This forms a valid chain and requires no recombinations. Thus, the total score is (j − 1)d + 2d + (n − j)d = 2d + (n − 1)d.
If a chain exists with score 2d + (n − 1)d, then in this chain, no recombinations are used, and anchors starting at all the anchor starting positions in Q are used.
For a given x, only one anchor to either vx,0 or vx,1 can be used while maintaining the precedence condition. Because each anchor to the center of G has a weight of two, the total contribution of the center of G to the score is at most 2d. The remaining anchors in the chain have a weight of one, and there are at most (n − 1)d of them after d anchors are used in the center. Hence, the maximum possible score is 2d + (n − 1)d. Notice also that taking d anchors in the center is the only way to achieve such a score.
The claim that there are no recombinations follows, as the maximum possible score using recombination becomes 2d + (n − 1)d − 1. The second claim follows because any chain using fewer than dn anchors cannot achieve the maximum score of 2d + (n − 1)d.
If a chain exists with the score 2d + (n − 1)d, then in this chain, all anchors for some vector gadget in Q are to the center of G.
If anchors from two different vector gadgets in Q are to the center of G, then because the center is contributing 2d to the score (the only way a score of 2d + (n − 1)d is achieved), there must be anchors M and 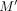 from two different vector gadgets that occur in adjacent vertices in the center, say with
from two different vector gadgets that occur in adjacent vertices in the center, say with 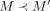 . However, this implies that there are at least d − 1 anchor positions in Q between M and
. However, this implies that there are at least d − 1 anchor positions in Q between M and 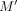 are not used in the solution, contradicting Lemma 9.
are not used in the solution, contradicting Lemma 9.
If there exists a chain with score 2d + (n − 1)d, then there exists orthogonal vectors ai and bj.
By Lemma 10, some vector gadget VG(bj) has all of its anchors going to the center of G. By Lemma 9, no recombinations are used, and these must lie on some path for a haplotype constructed from a vector ai. By design of the component gadgets, for 1 ≤ x ≤ d if ai[x] = 1 we get an anchor between vx,1 on the haplotype path for ai and VG(bj) if and only if bj[x] = 0. We conclude that ai and bj are orthogonal.
Observe that the total number of Mleft anchors is dn, the total number of Mright anchors is dn, and the total number of Mcenter anchors is at most 2dn; thus, N = O(dn). The number of haplotypes is 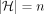 . Furthermore, |Q| = O(dn) and the graph G has O(d) vertices and edges, and the sum of vertex label lengths is O(dn). Hence, the reduction takes O(dn) time. Combined with Lemmas 8 and 11, it follows that an algorithm for Problem 4 running in time
. Furthermore, |Q| = O(dn) and the graph G has O(d) vertices and edges, and the sum of vertex label lengths is O(dn). Hence, the reduction takes O(dn) time. Combined with Lemmas 8 and 11, it follows that an algorithm for Problem 4 running in time 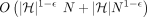 solves the OV problem in time
solves the OV problem in time 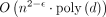 . This completes the proof of Lemma 7.
. This completes the proof of Lemma 7.
Problem 5 can be solved by extending Algorithm 1 (Supplemental Note S1). The extension requires a few additional terms corresponding to gap cost between adjacent anchors. This algorithm also runs
in 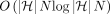 time. The reduction presented in Lemma 7 can be easily extended for Problem 5 as well.
time. The reduction presented in Lemma 7 can be easily extended for Problem 5 as well.
Implementation and benchmarking details
Software implementation
We implemented Algorithm 1 and the modifications to include gap penalty (for description, see Supplemental Note S1). We added our implementation in the Minichain code (Chandra and Jain 2023). Our new chaining implementation replaces the haplotype-agnostic chaining implementation proposed in our previous work.
Minichain supports an end-to-end seed-chain-extend workflow to align long reads and contigs to acyclic pangenome graphs. Similar
to the previous version of the software, we use the seeding and base-to-base alignment routines from Minigraph (Li 2018). We leave the implementation of our 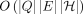 time base-to-base alignment algorithm (Methods) as future work because it requires further practical optimizations to reduce
the runtime, for example, using banded alignment or wavefront heuristics (Rautiainen and Marschall 2020; Marco-Sola et al. 2021; Zhang et al. 2022).
time base-to-base alignment algorithm (Methods) as future work because it requires further practical optimizations to reduce
the runtime, for example, using banded alignment or wavefront heuristics (Rautiainen and Marschall 2020; Marco-Sola et al. 2021; Zhang et al. 2022).
In Minichain, we parse the set of vertices, edges, and haplotype paths from a graphical fragment assembly (GFA) file (https://github.com/GFA-spec/GFA-spec). A read may be sampled from the same or the opposite orientation with respect to the graph. Accordingly, for each connected component in the input graph, we create another component of the same size with reverse-complemented vertex labels and reversed edges. We use the same path cover for the second component as the original component, but the direction of each path is reversed. We compute anchors by using Minigraph's seeding method, which finds (w, k)-minimizer matches (Roberts et al. 2004) between the query sequence Q and the vertex string labels σ(v) for all v ∈ V. The default values of w and k are 11 and 17, respectively. We set the weight of each anchor as 200 · k by default based on our experimental observations. Thus for k = 17, each anchor in a chain contributes 3400 to the total chain score. The multiplicative factor of 200 prevents gap penalties to dominate the overall chain score. Minichain can report multiple best-scoring chains and assign a mapping quality (Li et al. 2008) to each chain; the criteria for this remains same as before (Chandra and Jain 2023). We evaluate the chains produced by our algorithm without subjecting it to further Minigraph's heuristics.
Pangenome graph construction
We used 60 publicly available complete MHC haplotype sequences (https://zenodo.org/records/6617246). This set of 60 MHC haplotype sequences is composed of the CHM13 MHC haplotype (Nurk et al. 2022) and 59 other haplotypes from the Human Pangenome Reference Consortium (Liao et al. 2023). The length of these sequences varies from 4.8 Mbp to 5.1 Mbp. The MHC region is known to have significant polymorphism, inter-gene sequence similarity, and long-range haplotype structures (Dilthey 2021). We used Minigraph (v0.20-r559) (Li et al. 2020) to build an acyclic pangenome graph using the 60 MHC haplotypes. The graph construction algorithm in Minigraph augments structural variants (SVs) of size ≥50 bp in the graph. Our MHC pangenome graph contains 984 vertices, 1387 edges, and 210 SVs. Because Minigraph does not output haplotype paths in the graph, we realigned each haplotype to the graph to obtain the corresponding paths.
Simulation of query sequences
We simulated 135 MHC haplotype sequences. Each sequence was simulated as an imperfect mosaic of the 60 reference haplotypes (Fig. 5). We used the following procedure for simulation: (1) select a random haplotype path in the graph; (2) read the first l bases from the chosen haplotype path in the graph; (3) if the (l + 1)th base is in vertex v and |haps(v)| ≥ 2, then switch to another randomly chosen haplotype path at vertex v; and (4) read the next l bases, and repeat the procedure until we hit the last base of a haplotype path. After generating a sequence, we added substitutions at randomly chosen loci in the sequence. These substitutions approximately mimic true mutations and sequencing errors in real data. For each substitution rate of 0.1%, 1%, and 5%, we simulated 45 sequences. In each case, three sets of 15 sequences were simulated using l = 1 Mbp, l = 2 Mbp, and l = 3 Mbp, respectively. Using this procedure, the number of recombination events in all the simulated sequences ranged from one to four. Our pangenome graph and the set of query sequences are available on Zenodo (https://doi.org/10.5281/zenodo.10665350).

An example to illustrate simulation of a query sequence as a mosaic of reference haplotypes. In this example, l = 19.
Results
Comparison with haplotype-agnostic chaining using simulated data
We demonstrate that integrating recombination penalty in the sequence-to-graph chaining problem formulation leads to an improved concordance of the haplotypes used in our chains and the ground truth. A positive finite recombination penalty in our problem formulation allows the user to limit the recombinations in an optimal chain. If we set recombination penalty γ = 0, then our algorithm is equivalent to the haplotype-agnostic chaining algorithm of Chandra and Jain (2023). Recombination penalty γ = ∞ corresponds to haplotype-restricted chaining, where a chain can use only one of the reference haplotypes. We evaluated Minichain by using simulated MHC haplotype sequences and different values of recombination penalty γ = 0, 103, 104, 105, 106, ∞. Minichain computes an optimal chain as a sequence of pairs of the selected anchors and the haplotype indices. Thus, we know the sequence of haplotype indices and the number of recombinations in each chain.
We show the Pearson correlation coefficients between the observed number of recombinations and the true number of recombinations using different values of substitution rates and recombination penalties (Fig. 6A–C). The results suggest that γ = 104 gives the best correlation across different substitution rates. The correlation is weak when γ = 0 or γ = 106. The correlation is not defined when γ = ∞ because the observed number of recombinations remained zero; thus, the standard deviation of the observed values remained zero. In the haplotype-agnostic setting (γ = 0), a highest-scoring chain of minimizers can contain incorrect haplotype switches (for an example, see Supplemental Fig. S1). γ = 103 is also too low for setting recombination penalty because a single anchor in a chain contributes 3400 to the chaining score (Methods), and the algorithm may still favor an additional anchor even if it involves an incorrect haplotype switch. Using γ ≥ 104, the recombination penalty equals the score contributed by three or more anchors.
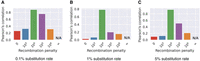
Pearson's correlation between the number of recombinations in Minichain's output chain and the true count. We evaluated the performance with substitution rates 0.1% (A), 1% (B), and 5% (C) and different recombination penalties.
Next, we compared the list of query sequences’ source haplotypes to the selected haplotypes in the output chains. Suppose
a query sequence was simulated using haplotypes in the order 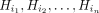 ; then, we recorded the “true haplotype recombination pairs” as a set of pairs
; then, we recorded the “true haplotype recombination pairs” as a set of pairs 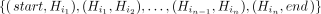 . Similarly, we recorded the sets corresponding to the “observed haplotype recombination pairs” from Minichain's output chains.
We compared these two sets for each query and calculated the F1-scores (Fig. 7A–C; Supplemental Fig. S4; Supplemental Table S1). In this experiment, precision was determined by the fraction of observed recombination pairs that were in the true set.
Similarly, recall was determined by the fraction of true recombination pairs that was observed in the output. We achieved
higher F1-scores with γ = 104, 105 compared with the haplotype-agnostic (γ = 0) and haplotype-restricted (γ = ∞) modes. These results suggest that haplotype-aware
chaining and alignment algorithms can be useful to prevent spurious recombinations.
. Similarly, we recorded the sets corresponding to the “observed haplotype recombination pairs” from Minichain's output chains.
We compared these two sets for each query and calculated the F1-scores (Fig. 7A–C; Supplemental Fig. S4; Supplemental Table S1). In this experiment, precision was determined by the fraction of observed recombination pairs that were in the true set.
Similarly, recall was determined by the fraction of true recombination pairs that was observed in the output. We achieved
higher F1-scores with γ = 104, 105 compared with the haplotype-agnostic (γ = 0) and haplotype-restricted (γ = ∞) modes. These results suggest that haplotype-aware
chaining and alignment algorithms can be useful to prevent spurious recombinations.

Box plots show the levels of consistency between the haplotype recombination pairs in Minichain's output chain and the ground truth using three different sets of simulated MHC sequences with substitution rates 0.1% (A), 1% (B), and 5% (C). We tested using different recombination penalties. Each red dot in the plots corresponds to a query sequence. The median values are highlighted in green.
We restricted the maximum count of recombinations in the simulated MHC haplotypes to four in the above experiments because MHC sequences are known to have long-range haplotype structures (Dilthey 2021). We performed additional experiments with higher number of recombination events (range: three to nine) and reached similar conclusions (Supplemental Figs. S2, S3).
Our MHC pangenome graph comprises SVs only. We expect further improvements in the accuracy of our algorithm if substitution and short indel variation are also augmented in a pangenome graph (Hickey et al. 2024). This is because more variants will help to distinguish between near-identical or closely related haplotypes. Doing this will also require improvements in Minichain's seeding and chaining implementation to allow the use of anchors that cover small bubbles in a graph. This is possible by considering a more flexible definition of anchors that can span multiple vertices (Rossi et al. 2022; Rizzo et al. 2023a,b). Our implementation uses minimizers, which also have a known weakness of being context dependent; that is, the decision to sample a k-mer is made based on its neighboring k-mers (Edgar 2021).
The runtime of Minichain ranged from 5 to 11 min for aligning a simulated MHC sequence to the pangenome graph. The worst-case time complexity of the proposed haplotype-aware chaining algorithm is higher compared with the existing haplotype-agnostic chaining algorithms. This is because we require haplotype paths as the path cover, whereas the haplotype-agnostic chaining algorithms use a path cover of minimum size (Mäkinen et al. 2019; Chandra and Jain 2023; Ma et al. 2023). For example, our graph with 60 haplotype paths has a minimum path cover of size five. Thus, we observed that our haplotype-aware chaining algorithm in Minichain was about 10× slower compared to the haplotype-agnostic chaining algorithm (Chandra and Jain 2023). Space requirements of our algorithm are also proportional to the count of haplotype paths (e.g., to store the score table C). As a result, we observed that the haplotype-aware algorithm required about 15× more memory compared with the haplotype-agnostic algorithm (Chandra and Jain 2023).
Evaluation using real data
We evaluated Minichain by using real long-read sequencing data sets from CHM13 human genome. Our MHC pangenome graph includes CHM13 as one of the 60 haplotypes. Because the CHM13 genome is effectively haploid, we expect all chains to be consistent with the CHM13 haplotype path in the graph. We used two PacBio HiFi data sets (obtained from the NCBI Sequence Read Archive [SRA; https://www.ncbi.nlm.nih.gov/sra] under accession numbers SRX5633451, SRR11292121) and one ONT data set (SRA; SRR23365080). We filtered the reads sampled from MHC region by mapping the reads to T2T-CHM13v2.0 genome reference (GCF_009914755.1) using minimap2 v2.26 (Li 2018). Next, we discarded reads of length ≤1 kbp. After these filtering steps, the N50 read lengths of the three data sets were 11 kbp, 20 kbp, and 83 kbp, respectively. We considered the chaining output of a read as correct if all the anchors in the highest-scoring chain overlapped with the CHM13 haplotype path in the pangenome graph.
The absolute fractions of correctly chained reads using the haplotype-agnostic objective (γ = 0) on the three data sets were 97.3%, 96.1%, and 91.1%, respectively. Using recombination penalty γ = 105, the fraction improved to 97.8%, 97.2%, and 94.2% respectively. Relatively speaking, the fraction of incorrect chains were reduced by 19%, 28%, and 35%, respectively.
These results, although preliminary, suggest that the proposed haplotype-aware sequence-to-graph alignment algorithms can be useful for genotyping and variant calling using pangenome references.
Discussion
Pangenome graph has emerged as a powerful alternative representation as a reference during resequencing of a genome. These graphs can be represented in either a haplotype-agnostic or haplotype-aware manner. The former excludes the information of the reference haplotypes, whereas the latter incorporates it. Leveraging the haplotype path information during read-to-graph or contig-to-graph mapping can be useful to compute accurate alignments and avoid unlikely haplotype recombinations. This will become even more important in the near future as more reference-quality human genomes are produced and include in a human pangenome graph. Prior work on haplotype-aware mapping has focused on restricting the recombination count to either zero or one (Mokveld et al. 2020; Sirén et al. 2020, 2021).
In this paper, we showed how the routine chaining and alignment tasks can be rigorously formulated and solved in a haplotype-aware manner. Our formulations focus on optimizing the standard metrics, for example, edit distance for alignment and chaining score for colinear chaining, while considering the switches used between the reference haplotypes. Thus, the proposed algorithms will help address the combinatorial challenges as more variants are included in a graph. The other key theoretical contribution is that we proved that our alignment and chaining algorithms are near-optimal assuming SETH. Subsequently, we demonstrated the practical value of our haplotype-aware chaining algorithm. We developed a proof-of-concept implementation in Minichain (v1.3). Our experiments show that the algorithm produces alignment with fewer false recombinations and better selection of the correct haplotypes.
More work is needed, especially to establish that the proposed haplotype-aware mapping algorithms can lead to improvements in routine genomic applications. This requires a more careful engineering of Minichain to make it useful for graphs that comprise SNPs and indels. The current implementation of Minichain is restricted to acyclic pangenome graphs that contain structural variation only. Lastly, it will also be interesting to investigate whether the genotype imputation accuracy can be improved using the proposed algorithms on low-pass whole-genome sequencing data.
Software availability
The Minichain aligner is available at GitHub (https://github.com/at-cg/minichain) and as Supplemental Code. The scripts and data sets to reproduce the results are available at GitHub (https://github.com/at-cg/minichain/tree/master/data/v1.3) and Zenodo (https://doi.org/10.5281/zenodo.10665350), respectively.
Competing interest statement
The authors declare no competing interests.
Acknowledgments
This work is supported by funding from the National Supercomputing Mission India, DBT/Wellcome Trust India Alliance Fellowship (grant no. IA/I/23/2/506979), and the Intel India Research Fellowship. We used computing resources at the C-DAC National PARAM Supercomputing Facility and the U.S. National Energy Research Scientific Computing Center.
Author contributions: All authors contributed to the development of the methods and proofs. G.C. implemented the software and performed the experiments.
Footnotes
-
[Supplemental material is available for this article.]
-
Article published online before print. Article, supplemental material, and publication date are at https://www.genome.org/cgi/doi/10.1101/gr.279143.124.
- Received February 15, 2024.
- Accepted June 24, 2024.
This article is distributed exclusively by Cold Spring Harbor Laboratory Press for the first six months after the full-issue publication date (see https://genome.cshlp.org/site/misc/terms.xhtml). After six months, it is available under a Creative Commons License (Attribution-NonCommercial 4.0 International), as described at http://creativecommons.org/licenses/by-nc/4.0/.








