Abstract
Genome evolution entails changes in the DNA sequence of genes and intergenic regions, changes in gene numbers, and also changes in gene order along the chromosomes. Genes are reshuffled by chromosomal rearrangements such as deletions/insertions, inversions, translocations, and transpositions. Here we report a comparative study of genome organization in the main African malaria vector,Anopheles gambiae, relative to the recently determined sequence of the Drosophila melanogaster genome. The ancestral lines of these two dipteran insects are thought to have separated ∼250 Myr, a long period that makes this genome comparison especially interesting. Sequence comparisons have identified 113 pairs of putative orthologs of the two species. Chromosomal mapping of orthologous genes reveals that each polytene chromosome arm has a homolog in the other species. Between 41% and 73% of the known orthologous genes remain linked in the respective homologous chromosomal arms, with the remainder translocated to various nonhomologous arms. Within homologous arms, gene order is extensively reshuffled, but a limited degree of conserved local synteny (microsynteny) can be recognized.
Modern genomics have revolutionized genetics and, consequently, biology. The enormous acceleration of data acquisition, in fields such as whole genome sequence determination and genome-wide gene expression profiling, has opened novel possibilities for the study of model organisms and organisms for which, until recently, only rudimentary biological knowledge was available (orphan organisms). For example, until a decade ago only a few tens of genes had been identified in important insect disease vectors such asAnopheles gambiae or Aedes aegypti, which now number ∼24,000 and 1700 entries, respectively, in the nucleic acid databases. Many of these represent partial genomic sequences, sequence tagged sites (STSs), and anonymous cDNAs or expressed sequence tags (ESTs; for review, see Louis 1999). Such genetic and molecular genetic information may prove helpful in designing new schemes to fight the diseases transmitted by these mosquitoes, such as malaria and dengue (James et al. 1999). Progress in elucidating the genomic information of formerly orphan insect organisms can be considerably accelerated by using the closest available model organism, in this case Drosophila melanogaster, as a guide.
A. gambiae s.s. (sensu stricto) is a member of the AfricanA. gambiae species complex that consists of six distinct sibling species and itself can be distinguished into a series of taxa or incipient species (Coluzzi et al. 1985), all differing in vectorial capacity (see Touré et al. 1998). The pioneering studies of Coluzzi and his collaborators on the construction of polytene maps for this species complex and the documentation of both fixed and polymorphic inversions can be viewed as the start of genomic research on the malaria mosquito.
Over the past decade, knowledge of the molecular biology and genetics of A. gambiae s.s. has vastly improved. For example, numerous molecular studies of the interactions between Anopheles andPlasmodium have radically improved our understanding of this vector-parasite system (for review, see Sinden 1999). The molecular study of the genome was initiated with the construction of a first low-resolution physical map, linked to the polytene chromosomes (Zheng et al. 1991), followed by the construction of a detailed, microsatellite-based recombination map (Zheng et al. 1993, 1996). Integration of the genetic (recombinational), cytogenetic (polytene), and molecular (clone and sequence) maps has progressed rapidly; it entails the genetic and cytogenetic mapping of random amplified polymorphic DNA (RAPD) markers (Dimopoulos et al. 1996a), the recombinational mapping of microsatellites, and the assignment of both microsatellites and anonymous DNA markers to specific chromosomal locations, using in situ hybridization to polytene chromosomes (della Torre et al. 1996; Dimopoulos et al. 1996a; Zheng et al. 1996; Wang et al. 1999). Microsatellites have been used successfully both for gene mapping (Collins et al. 1997; Zheng et al. 1997; Ranson et al. 2000) and for studies of population biology (e.g., see Lanzaro et al. 1998;Kamau et al. 1999; Wang et al. 1999, 2001). Finally, routine germline transformation and thus reverse genetic studies of A. gambiaecan be expected soon, judging by the recent success in transforming both anopheline (A. stephensi; Catteruccia et al. 2000) and aedine mosquitoes (Ae. aegypti; Coates et al. 1998;Jasinskiene et al. 1998).
Important additional tools for comparative genomic studies of A. gambiae have become available recently. They include a collection of ESTs that may represent ∼10% of the mosquito genes (Dimopoulos et al. 2000), and ∼17,500 sequence-tagged ends of a bacterial artificial chromosomes (BAC) chromosomal library representing 14.5 Mb or 7% of the expected euchromatic DNA sequence (http://bioweb.pasteur.fr/BBMI; C. Roth and F.H. Collins, pers. comm.). An experimental strategy that combines the identification of orthologs by sequence similarity searches and their mapping to the chromosomes or linkage groups of different species has proven to be very informative in comparative genomic studies of both animals (O'Brien et al. 1999) and plants (Terryn et al. 1999). An important type of information derived from such studies is the degree of conserved synteny: to what extent the chromosomal dynamics in evolution permit linkage group conservation, that is, persistent linkage of most genes in a given chromosome between compared species (long-range synteny in homologous chromosomes). A second important issue is to what extent originally neighboring genes remain clustered (local conserved synteny, or microsynteny) rather than becoming randomized in terms of their order within the homologous chromosome.
Here we used the essentially complete sequence information on theD. melanogaster genome (Adams et al. 2000), together with the available A. gambiae genomic resources, to address the questions of sequence conservation, long-range synteny, and local microsynteny between the genomes of the mosquito and the fruit fly, two distantly related diptera.
RESULTS
Chromosomal Distribution of A. gambiae Orthologs of Genes From Two D. melanogaster Chromosomal Regions
In a first set of experiments aiming at exploring long-range synteny and microsynteny, we identified, among the currently available A. gambiae sequences, putative orthologs of genes in which in D. melanogaster are clustered within two well-studied chromosomal regions, each nearly 3 Mb long. We then determined the genomic locations of these putative orthologs by hybridization to the mosquito polytene chromosomes.
The fruit fly genomic regions that were chosen for these experiments have been completely sequenced and annotated, both in clone-by-clone sequencing projects and as part of whole-genome shotgun sequencing. One of these Drosophila regions is the autosomal Adhregion, covering 2.9 Mb on both sides of the Adh gene, in divisions 34B–35F of chromosomal arm 2L (Ashburner et al. 1999). The other is the tip of chromosome X, encompassing 2.6 Mb in polytene divisions 1–3 (Benos et al. 2000,2001). Both of these regions were also covered by whole-genome shotgun sequencing (Adams et al. 2000).
The 256 genes from the tip of the X and the 219 genes from theAdh region of Drosophila were used to query, byTBLASTN, collections of both STSs and ESTs ofAnopheles: the 17,506 STSs representing end sequences of BAC clones, and the 6012 ESTs that correspond to 2380 potential genes (cDNA clone clusters from a subtracted normalized library; Dimopoulos et al. 2000). To define genes as putative orthologs, hits that satisfied criteria of high score of >40, probability P(N) of <1 e−10, and percentage of identical amino acid residues >30 over a long range were selected in a first round. From them, all spurious hits that were caused by the presence of low complexity segments were eliminated, and the remaining hits were confirmed byBLASTX analysis as best bidirectional hits against a database of 14,080 amino acid sequences of known and predictedDrosophila genes (release 1.0; Adams et al. 2000). Those that passed this test were further verified by direct comparison to the corresponding Drosophila entry, taking into account potential intron-exon boundaries. Henceforth, these validated genes will be referred to as orthologs for convenience (see also Discussion). These procedures (see also Methods) identified 19 mosquito orthologs of unique genes found in the tip region of the Drosophila Xchromosome and 31 orthologs of unique genes found in theDrosophila Adh region. For greater accuracy, we eliminated from consideration additional probable orthologs (18 showing hits toX-tip and nine showing hits to Adh region genes), because they belong to chromosomally dispersed multigene families. This was necessary because the true ortholog can not be chosen among the different members of a given gene family until both genomes are fully sequenced.
The 50 orthologs that were retained for further analysis were present in 33 BAC and 37 cDNA clones (a number of them were detected by both STSs and ESTs). Representative clones were used as probes for in situ hybridization analysis to A. gambiae polytene chromosomes. Tables 1 and 2include the results of this analysis for the X-tip orthologs and Adh region orthologs, respectively. Notably, the Tables show cytogenetic and molecular locations of the 50 Drosophilagenes and the sequence identifiers and cytogenetic locations of the corresponding A. gambiae orthologs. The distribution of theX-Tip and Adh region orthologs among the five polytene chromosome arms of A. gambiae are tabulated in Table3, together with the results of statistical analysis of these distributions using the binomial test of significance, confirmed by the χ2 test.
Cytological Mapping of Putative Anopheles gambiaeOrthologues of Genes in the Drosophila melanogasterDivisions 1–3 of Chromosome X
| D. melanogaster | % Identical amino acids | A. gambiae | ||||
| Cytology | Mol. map | Gene | Sequence | Accession no. | Cytology | |
| 1B4-5 | 263 | svr | 61 | 4A3B-AAA-E-12-R | AJ284634 | 24:12B |
| 1B5-7 | 294 | EG:65F1.1 | 34–46 | 02A19-t7 | AL140406 | 2L:21C |
| 1B10 | 394 | RpL36 | 83 | 4A3A-P7A4-F | AJ281575 | 3R:32A |
| 1B10-11 | 412 | EG:115C2.11 | 97 | 4A3B-AAX-A-12-R | AJ285778 | 2L:21F |
| 1B10-11 | 420 | spkA | 92 | 4A3B-AAI-C-06-F | AJ284038 | 3R:35C |
| 1B12-C1 | 451 | sdk | 58–78 | 06M18-sp6 | AL143155 | X:1D |
| 1C5-D1 | 704 | EG:BACR7A4.5 | 48–62 | 21I19-sp6 | AL154783 | 3L:38B |
| 1D4-E1 | 797 | EG:BACR7A4.8 | 51 | 4A3A-AAT-H-11-R | AJ282437 | 3L:44D |
| 2A2-3 | 1137 | EG:132E8.3 | 78 | 4A3A-P3A1-F | AJ281385 | 2R:15A |
| 2A4-B1 | 1188 | EG:49E4.1 | 90 | 25H04-sp6 | AL153026 | 2R:7A |
| 2B1-2 | 1239 | sta | 75 | 4A3A-AAT-D-12-R | AJ282394 | 3L:44B |
| 2B6-7 | 1417 | EG:171E4.4 | 48 | 21P07-t7 | AL151067 | X:2C |
| 2B9-10 | 1521 | Adar | 49–87 | 27D16-sp6 | AL154010 | X:4A |
| 2B13-14 | 1621 | EG:63B12.4 | 61 | 4A3A-AAT-B-06-R | AJ282366 | 2R:12C |
| 2B13-15 | 1637 | trr | 45–95 | 32G18-sp6 | AL157001 | 3L:41D |
| 2B14-16 | 1648 | arm | 76 | 4A3A-AAQ-E-05-F | AJ280532 | multiple |
| 2D4-E1 | 1893 | Pgd | 29–68 | 30A12-sp6 | AL155664 | 3R:31D |
| 2F1-2 | 2022 | EG:25E8.1 | 67 | 4A3A-ABB-H-01-F | AJ281139 | 2R:9B |
| 2F3 | 2077 | EG:BACH48C10.2 | 40 | 4A3B-AAW-F-07-R | AJ285740 | 3R:33B |
[i] For D. melanogaster, gene names and cytological localization are derived from FlyBase; the genes are ordered according to the molecular map, which represents the approximate positions (in kilobases) of the 5′ ends of the genes (Benos et al. 2001). ForA. gambiae, the cytological localizations on polytene chromosomes are according tot he maps of M. Coluzzi, A. Sabbatini, M.A. Di Deco, and V. Petrarca (pers. comm.); the names of the sequences are as they appear in corresponding database entries with the indicated accession numbers. The A. gambiae 4A3A and 4A3B entries are expressed sequence tags, and entries ending with sp6 or t7 are sequence tagged sites (STSs) from chromosomal bacterial artificial chromosomes (BACs), % identities refer to one or more exons.
Cytological Mapping of Putative Anopheles gambiaeorthologs of Genes in the Drosophila melanogaster AdhRegion of Chromosome 2L
| D. melanogaster | % Identical amino acids | A. gambiae | ||||
| Cytology | Mol. map | Gene | Sequence | Accession no. | Cytology | |
| 34B11-C1 | 44 | B4 | 46–76 | 31F12-sp6 | AL156399 | 3R:29A |
| 34C3-4 | 130 | BG:BACR48E02.4 | 79 | 4A3B-AAC-G-09-R | AJ284828 | 3R:31C(1) |
| 34D1 | 294 | BG:DS00797.7 | 80 | 4A3A-AAK-H-12-F | AJ280101 | 3R:32B(2) |
| 34D1-3 | 306 | adat | 48–53 | 05M01-sp6 | AL142526 | 3R:33C(3) |
| 34D1-3 | 316 | b | 47–63 | 12K15-sp6 | AL146601 | 3R:32B(2) |
| 34D2-4 | 324 | Spo2 | 88 | 4A3A-ABA-B-10-R | AJ282629 | 3R:32B(2) |
| 34D3-4 | 328 | Rpll33 | 84 | 4A3B-AAC-H-08-R | AJ284838 | 3L:38A |
| 34E2-4 | 508 | bgm | 55–76 | 31H08-sp6 | AL156466 | 3R:30E |
| 34F1-2 | 728 | smi35A | 82 | 26G10-t7 | AL153535 | 3R:29B |
| 34F5-A1 | 848 | Rab14 | 66–95 | 32D17-sp6 | AL156885 | 3R:30D |
| 35A1 | 853 | I(2)35Aa | 57 | 4A3A-AAN-C-07-R | AJ281979 | 2R:19B |
| 35A1 | 857 | spel1 | 60–75 | 25H11-sp6 | AL153035 | 3R:37D |
| 35B8 | 1433 | BG:SD01219.3 | 65 | 23G11-sp6 | AL151883 | 3R:35B |
| 35B9 | 1498 | BG:DS00929.3 | 88 | 4A3B-AAE-D-12-R | AJ284976 | 3L:38A |
| 35C1 | 1548 | ck | 46–60 | 27M17-sp6 | AL154364 | 3R:30C |
| 35CD1-2 | 1599 | BG:DS04929.3 | 51–85 | 02O08-sp6 | AL140935 | 3R:36C |
| 35D3 | 1975 | lace | 58–79 | 11J20-t7 | AL146045 | 3R:29C(4) |
| 35D4 | 2146 | BG:DS07108.2 | 58–72 | 16K02-sp6 | AL148101 | 3R:34D |
| 35D4 | 2185 | CycE | 54–70 | 24K22-sp6 | AL152578 | 3R:30A |
| 35E1-2 | 2411 | beat-B | 49–77 | 03I12-t7 | AL141289 | 3R:33C(3) |
| 35F1 | 2500 | beat | 62–69 | 03I12-sp6 | AL141288 | 3R:33C(3) |
| 35F6-8 | 2739 | BG:DS02740.4 | 55 | 4A3A-ABC-D-11-F | AJ281189 | 3R:29C(4) |
| 35F7-8 | 2746 | Cyp303a1 | 51 | 09O07-t7 | AL145116 | 3R:36D-37A |
| 35F7-8 | 2750 | heix | 70 | 4A3A-ABA-E-06-F | AJ281029 | 2R:17A |
| 35F7-8 | 2757 | CG5861 | 78 | 4A3B-AAX-D-01-F | AJ284498 | 3R:31C(1) |
| 35F7-8 | 2759 | Sed5 | 83 | 4A3B-AAF-G-04-R | AJ285095 | 3R:31C(1) |
| 35F7-8 | 2761 | cni | 82 | 4A3A-AAY-B-08-R | AJ282455 | 2R:9A |
| 35F8-9 | 2776 | cact | 51 | 4A3A-AAT-A-11-R | AJ282359 | 3R:29C(4) |
| 35F8-9 | 2780 | I(2)35Fe | 68 | 4A3A-AAS-G-11-R | AJ282339 | 3R:29C(4) |
| 35F9-11 | 2807 | BG:DS09218.4 | 63–78 | 27H11-t7 | AL154163 | 2R:9C |
| 35F10-11 | 2816 | BG:DS09218.5 | 51–58 | 32O04-t7 | AL157296 | 2L:21F |
[i] Legend as in Table 1. Underlined Drosophila genes show transposition to chromosomes of A. gambiae other than3R. The superscripts 1–4 mark clones that are co-localized within subdivision 31C, 32B, 33C, and 29C, respectively (see Results).
Chromosomal Distribution of A. gambiae Orthologs ofD. melanogaster Genes
| Drosophilaregions, no. of observed and expected genes, and P values of binomial test | A. gambiae polytene chromosome arms | ||||
| X | 2R | 2L | 3R | 3L | |
| X-tip, divisions 1–3 (19 genes) | |||||
| Observed genes | 3 | 5 | 2 | 4 | 5 |
| Expected genes | 2.27 | 5.58 | 4.13 | 3.82 | 3.20 |
| P value of binomial test | 0.719 | 0.81 | 0.283 | 1 | 0.35 |
| Adh region of2L (31 genes) | |||||
| Observed genes | 0 | 4 | 1 | 24 | 2 |
| Expected genes | 3.71 | 9.10 | 6.74 | 6.23 | 5.22 |
| P value of binomial test | (0.028) | (0.048) | (0.0079) | 9.939 e−12 | 0.15 |
[i] The binomial test was applied as follows. For each of the twoD.melanogaster regions analyzed, the number of putative orthologs observed in each A. gambiae polytene chromosome arm was listed. The corresponding expected number of orthologs was calculated according to the number of lettered subdivisions perAnopheles chromosomal arm (see Results), and the P value was determined by the binomial test. Statistically significant results are shown in bold, within or without parantheses depending on whether genes are underrepresented or overrepresented in that arm, respectively. Note that Adh region orthologs are significantly overrepresented in the mosquito 3R arm and underrepresented in X, 2R,and 2L.
For the statistical analysis, we compared the number of orthologs corresponding to each Drosophila region that were observed in each chromosomal arm of the mosquito to the number expected if the association were random according to chromosomal arm length. To calculate the expected numbers, the lengths of the five mosquito chromosomal arms were estimated according to the number of their lettered subdivisions, as recognized in the map of Coluzzi and associates (22 subdivisions for X, 54 for 2R, 40 for2L, 37 for 3R, and 31 for 3L; or 12.0%, 29.35%, 21.7%, 20.1%, and 16.85% of the total, respectively; the map is accessible at http://www.anodb.gr/AnoDB/Cytomap/). The binomial test is an exact probability test that is used to examine the distribution of a single dichotomy in conditions when only a relatively small sample is available, as is the case here. It provides a one-sample test of the difference between the sampled distribution and a given distribution. In this case, the given distribution is based on the null hypothesis that the genes of each Drosophilachromosomal region are randomly redistributed across all five chromosome arms of A. gambiae according to their lengths. As shown in Table 3 for the gene probes derived from the tip of chromosomeX, all P values are >0.05, and thus, the null hypothesis cannot be rejected. Similarly, the χ2 statistic (not shown) is equal to 2.415, lower than the critical value χ2 4[0.05] = 9.49; therefore, the null hypothesis can not be rejected. By these criteria, none of the five mosquito chromosomal arms is significantly enriched for orthologs of the X-tip genes of Drosophila.
In contrast, the results of the same analysis strongly indicate that the Drosophila arm 2L (at least its Adhregion) corresponds to the chromosome arm 3R of A. gambiae (P = 9.939 e−12). That mosquito arm includes nearly fourfold as many genes as expected: It contains 24 (77%) of the currently available orthologs of the Drosophila Adh region genes, whereas only 7 (23%) orthologs are scattered over three other mosquito autosomal arms. Furthermore, in three out of four remaining mosquito chromosomal arms (2R, 2L, and3L), the prevalence of orthologs of Drosophila 2Lgenes is statistically significantly lower than expected. Thus, the binomial test clearly rejects the null hypothesis of random redistribution of Adh region genes, in terms of both positive and negative correlations. Rejection is also supported by the χ2 analysis, in which the statistic (not shown) is equal to 64.12 with the same critical value as before (χ2 4[0.05] = 9.49).
It should be noted from Table 3 that the mosquito orthologs of theAdh region genes are not evenly distributed within the mosquito 3R arm: Half of them are located within four chromosomal subdivisions (29C, 31C, 32B, and 33A), whereas the other half are scattered among the other 33 subdivisions of 3R. This apparent clustering may correspond to microsynteny, as will be discussed below.
Distribution of Randomly Selected A. gambiae Sequences and Their D. melanogaster Orthologs
A similar but reverse method was used in a second experiment addressing the question of long-range synteny. In this case, we started by mapping random A. gambiae STSs mapped on the polytene chromosomes and determined their orthologs and the respective cytogenetic locations in D. melanogaster.
Randomly selected BAC clones of A. gambiae from the library that had been used to determine STS end sequences (http://bioweb.pasteur.fr/BBMI; C. Roth and F.H. Collins, pers. comm.) were mapped by in situ hybridization to mosquito polytene chromosomes. A total of 1217 STS were available from 720 cytogenetically mapped clones, and they were used for a BLASTX search of the protein sequences corresponding to the 14,080 known and predictedD. melanogaster genes (release 1.0; Adams et al. 2000). This search led to the identification of 49 mapped STS that were putative orthologs of unique D. melanogaster genes. In addition,A. gambiae genes of known cytogenetic location were used to search the same Drosophila database, yielding 21 additional hits. This number also included cecropin and ADP/ATP, two A. gambiae genes, each of which is homologous to a corresponding small multigene family in Drosophila, clustered at a single cytogenetic location. Table4 lists these 70 mosquito gene sequences by cytogenetic location, together with theirDrosophila orthologs and their locations. Table5 summarizes and correlates the chromosomal locations of corresponding sequences in the two species. As in the previous experiment, the binomial test and the confirming χ2 analysis (not shown) used the numbers of orthologs expected on each Drosophila chromosomal arm, in this case according to a random distribution calculated on the basis of the respective known DNA content of the Drosophila arms (Adams et al. 2000).
Cytological Position of Randomly Selected A. gambiaeSequences and Their D. melanogaster Orthologs With Unique Localization in the Genome
| A. gambiae | % Identical amino acids | D. melanogaster | ||||
| Cytology | Sequence/gene | Accession no. | Gene | Cytology | Reference | |
| X: 1C | cec | AF200686 | 45–52 | CecA1 | 3R:99E3-4 | Vizioll et al. 2000 |
| CecA2 | 3R:99E3-4 | |||||
| CecB | 3R:99E3-4 | |||||
| CecC | 3R: 99E3-4 | |||||
| X: 1D | 06M18-sp6 | AL143155 | 58 | sdk | X: 1B12 | This study |
| X: 2A | w | U29484 | 54–72 | w | X:3C2 | Besansky et al. 1995 |
| X: 2C | 21P07-t7 | AL151067 | 45 | EG: 171E4.4 | X:2B6-7 | This study |
| X: 3D | 20I10-t7 | AL150270 | 47–77 | CG1472 | 2R:46C1-4 | This study |
| X: 3D-4A | pKM42 | U50467 | 92 | Cycbeta100B | 3R:100B5-8 | della Torre et al. 1996 |
| X: 4A | 27D16-sp6 | AL154010 | 87 | Adar | X: 2B9 | This study |
| X: 4B | 10A05-sp6 | AL145180 | 43–75 | CG11428 | X:12A7-8 | This study |
| X: 4C | 10F12-sp6 | AL145357 | 36–80 | ey | 4:102D5 | This study |
| X: 5B | 10I08-t7 | AL145443 | 87–97 | sqh | X: 5D6 | This study |
| X: 5D | 10A22-t7 | AL145201 | 36 | CG15054 | X:17B1 | This study |
| X: 5D | 10L08-sp6 | AL145544 | 58 | CG6803 | 3R:88E5 | This study |
| X: 6 | 17N20-sp6 | AL148805 | 32–46 | CG8772 | 2R:49B8-9 | This study |
| 2R: 8C | 17I13-sp6 | AL148612 | 44–61 | CG7593 | 3R:99B11-C1 | This study |
| 2R: 11B | 30E10-t7 | AL155810 | 74 | ESTS:149B10S | 3L:66A8-10 | This study |
| 2R: 12A-B | pKM50 | U50471 | 66 | CG7808 | 3R:99C6-7 | della Torre et al. 1996 |
| 2R: 12C | 17M11-t7 | AL148757 | 39 | mod(r) | X:1B9-10 | This study |
| 2R: 14D | 10G21-t7 | AL145401 | 88 | CG9492 | 3L:70E3 | This study |
| 2R: 14E | pKM52 | U50472 | 57 | CG6783 | 3R:86E13-15 | della Torre et al. 1996 |
| 2R: 16A | 10D21-t7 | AL145303 | 70–73 | slo | 3R:96A18-20 | This study |
| 2R: 19B-E | Scr | AF080564 | 35–54 | Scr | 3R:84A5-B1 | Devenport et al. 2000 |
| 2R: 19D | 17P11-t7 | AL148868 | 47 | ninaC | 2L:27F5-6 | This study |
| 2R: 19D | Antp | AF080565 | 53 | Antp | 3R:84B2-4 | Devenport et al. 2000 |
| 2R: 19D | Ubx | AF080562 | 62 | Ubx | 3R:89D6-E2 | Devenport et al. 2000 |
| 2R: 19E | abd-A | AF080566 | 61–67 | abd-A | 3R:89E3-4 | Devenport et al. 2000 |
| 2L: 20C | 05B09-t7 | AL142182 | 47 | EG:BACH7M4.1 | X:2F5 | This study |
| 2L; 20D | 10F01-sp6 | AL145340 | 26–53 | CG10173 | 3L:65A7-8 | This study |
| 2L: 21C | 02A19-t7 | AL140406 | 34 | EG:65F1.1 | X:1B4 | This study |
| 2L;21F | 32O04-t7 | AL157296 | 52 | BG:DS09218.5 | 2L:35F10 | This study |
| 2L: 22C | 21G04-t7 | AL150716 | 42 | CG18289 | 2R:41F3 | This study |
| 2L: 22D | 19D01-sp6 | AL149568 | 55 | CG3790 | 2R:49D2-4 | This study |
| 2L: 22D | 18E22-t7 | AL149075 | 66 | CG10483 | 3L:64F4-5 | This study |
| 2L: 22E | 16A02-t7 | AL147784 | 31 | AAF49108.1 | 3L:76D2-3 | This study |
| 2L: 23B | 21I15-t7 | AL150818 | 37 | RecQ4 gene | 3L: 66B11 | This study |
| 2L; 23C | pKM2 | U50477 | 70 | CG7991 | 3L: 62B1 | della Torre et al. 1996 |
| 2L: 23D | 07F24-t7 | AL143530 | 39 | CG13076 | 3L:72D1 | This study |
| 2L: 24C | 16L07-sp6 | AL148140 | 83 | CG12215 | 2R:46EF | This study |
| 2L: 26A | ADP/ATP | L11617 | 76–80 | sesB | X:9E4-7 | Beard et al. 1994 |
| Ant2 | X:9E4-7 | |||||
| 2L; 27A | 32J02-sp6 | AL157092 | 31–85 | Ptpmeg | 3L:61C1 | This study |
| 2L: 27A | 27H17-sp6 | AL154170 | 60 | EG:52C10.1 | 2R:54E9 | This study |
| 2L: 27D | pKM134 | U50479 | 90 | RpS13 | 2L: 29B2-3 | della Torre et al. 1996 |
| 2L: 28A | 10F06-t7 | AL145346 | 48–66 | CG15373 | X:16F3 | This study |
| 2L: 28C | IanB2 | AJ271193 | 60 | LanB2 | 3L:68B10 | Viachou et al. 2001 |
| 2L: 28D | 22L01-t7 | AL151486 | 41–55 | CG4324 | 2R:60A16-B1 | This study |
| 3R: 29B | pKM3 | U50480 | 43–72 | Sin3A | 2R:49B2-3 | della Torre et al. 1996 |
| 3R: 30C | 27M17-sp6 | AL154364 | 46–60 | ck | 2L:35C1 | This study |
| 3R: 31B | 10G09-sp6 | AL145389 | 84 | CG9313 | 2R:57B11-12 | This study |
| 3R: 33A | cc41 | AF002238 | 50–73 | CG17489 | 2L:40B-D | A. Cornel, pers. comm. |
| 3R: 33B | bd | AF042732 | 51–58 | CG10655 | 2L:37B11-13 | Romans et al. 1999 |
| 3R: 33B | TU37B2 | AF042732 | 76–96 | CG10470 | 2L:37B11-13 | Romans et al. 1999 |
| 3R: 33B | Dox | AF042732 | 74 | Dox-A2 | 2L:37B11-13 | Romans et al. 1999 |
| 3R: 33B | Ddc | AF063021 | 74–80 | Ddc | 2L: 37C1 | Romans et al. 1999 |
| 3R: 34A | 25P12-sp6 | AL153311 | 68–90 | Cbp53E | 2R:53E6-11 | This study |
| 3R: 34C | 10F08-t7 | AL145350 | 56 | CG18252 | 2L:25C8-9 | This study |
| 3R: 34C | Gambif1 | X95911 | 44–73 | dl | 2L:36C2-3 | Barillas-Mury et al. 1996 |
| 3R: 35C | 10B06-t7 | AL145213 | 36–48 | CG11003 | 3L:69E1 | This study |
| 3R: 38B | 2BI19-sp6 | AL154783 | 56 | EG:BACR7A4.5 | X:1C5-D1 | This study |
| 3L: 40B | 24C03-t7 | AL152258 | 62 | CG1698 | 2R:46B7 | This study |
| 3L: 40B | 17M09-sp6 | AL148752 | 41 | CG4464 | 3L:67A1-3 | This study |
| 3L: 40B | 32L10-t7 | AL157191 | 54 | CG3252 | X: 4F5 | This study |
| 3L: 41D | 20D20-sp6 | AL150137 | 41 | CG18289 | 2R:41F3 | This study |
| 3L: 42A | defensin | X93562 | 56 | Def | 2R:46D7-9 | Richman et al. 1996 |
| 3L: 42A | 26B23-t7 | AL153383 | 43 | CG8642 | 2R:44D2 | This study |
| 3L: 42A | 30G20-sp6 | AL155891 | 42–88 | CG3186 | 2R:60B2-3 | This study |
| 3L: 42B | 10D11-t7 | AL145288 | 28–71 | CG6501 | 2R:54D1 | This study |
| 3L: 43A | 20B11-t7 | AL150058 | 41 | snk | 3R: 87D11 | This study |
| 3L: 45C | 03G02-sp6 | AL141206 | 67 | CG2103 | 3R:63A1-2 | This study |
| 3L: 46A | 19L20-t7 | AL149862 | 44 | CG7169 | 3L:78E4-5 | This study |
| 3L: 46A | 28J05-sp6 | AL154795 | 53 | CG8706 | 2R:44B3 | This study |
| 3L: 46CD | 19N23-sp6 | AL149940 | 40 | CG14196 | X: 18B6-8 | This study |
[i] Legend as in Table 1. A. gambiae sequence names are as they appear in the corresponding references or database entries.
Chromosomal Distribution of Putative D. melanogasterOrthologs of A. gambiae Genes
| A. gambiae polytene chromosome arms, observed and expected genes, and P values of the binomial test | D. melanogaster polytene chromosome arms | |||||
| X | 2L | 2R | 3L | 3R | 4 | |
| X(13 genes) | ||||||
| Observed genes | 7 | 0 | 2 | 0 | 3 | 1 |
| Expected genes | 2.39 | 2.52 | 2.35 | 2.67 | 3.07 | 0.13 |
| Pvalue of the binomial test | 0.0036 | 0.0508 | 0.2844 | 0.0503 | 0.2547 | 0.1152 |
| 2R(12 genes) | ||||||
| Observed genes | 1 | 1 | 0 | 2 | 8 | 0 |
| Expected genes | 2.21 | 2.33 | 2.17 | 2.47 | 2.83 | 0.12 |
| Pvalue of the binomial test | 0.2351 | 0.2169 | 0.0913 | 0.2791 | 0.0016 | 0.8864 |
| 2L(19 genes) | ||||||
| Observed genes | 4 | 2 | 5 | 8 | 0 | 0 |
| Expected genes | 3.49 | 3.68 | 3.43 | 3.91 | 4.49 | 0.19 |
| Pvalue of the binomial test | 0.2101 | 0.1651 | 0.1373 | 0.0193 | (0.0060) | 0.8262 |
| 3R(13 genes) | ||||||
| Observed genes | 1 | 8 | 3 | 1 | 0 | 0 |
| Expected genes | 2.39 | 2.52 | 2.35 | 2.67 | 3.07 | 0.13 |
| Pvalue of the binomial test | 0.2089 | 0.0009 | 0.2300 | 0.1692 | (0.0298) | 0.8775 |
| 3L(13 genes) | ||||||
| Observed genes | 2 | 0 | 7 | 2 | 2 | 0 |
| Expected genes | 2.39 | 2.52 | 2.35 | 2.67 | 3.07 | 0.13 |
| Pvalue of the binomial test | 0.2822 | 0.0608 | 0.0033 | 0.2624 | 0.2238 | 0.8775 |
[i] Legend as in Table 3, except that expected numbers and Pvalues were calculated for D. melanogaster chromosome arms. The actual DNA content of the D. melanogaster chromosome arms (Adams et al. 2000) was used for the calculations.
The data from this second experiment (Table 5) completely confirm and extend the conclusions from the first experiment. They identify statistically significant and unique chromosomal arm homologies with the P values ranging from 0.0193 to 0.0009, as follows:X Ag/X Dm,2R Ag/3R Dm,2L Ag/3L Dm,3R Ag/2L Dm, and3L Ag/2R Dm. Except for these, no other pairs even approach statistical significance as homologs. However, the dot chromosome 4 of Drosophila does not exist in the mosquito, and the single known Anopheles homolog of a chromosome 4 gene is found on the mosquito Xchromosome. In this second experiment, as much as in the first, the relative order of orthologous genes within the corresponding chromosomal arms of the two species appeared to be scrambled. Again, however, some residual microsynteny was detected (see below). For an additional statistical analysis of the same data, we took as a starting point the chromosomal distribution of the Drosophila orthologs and compared the observed and expected distributions ofAnopheles genes; this inverse comparison corresponds to that of the first experiment. As shown in Table6, the inverse P values are all significant, convincingly confirming the chromosomal arm homologies established from Table 5.
Chromosomal Distribution of Putative A. gambiaeOrthologs of D. melanogaster Genes
| Drosophila chromosome arm | Total observed genes | A. gambiae homolog | Orthologs on homolog | Expected orthologs | % of genes on homolog | Expected percentage | Inverse Pvalue | Pvalue |
| X | 15 | X | 7 | 1.79 | 46.7 | 12.0 | 0.0008 | 0.0036 |
| 2L | 11 | 3R | 8 | 3.52 | 72.7 | 29.4 | 0.0063 | 0.0016 |
| 2R | 17 | 3L | 7 | 3.04 | 41.2 | 21.7 | 0.0037 | 0.0193 |
| 3L | 13 | 2L | 8 | 2.41 | 61.5 | 20.1 | 0.0005 | 0.0009 |
| 3R | 13 | 2R | 8 | 3.03 | 61.5 | 16.9 | 0.0168 | 0.0033 |
[i] The binomial test was applied as follows. For each D. melanogaster chromosome arm, the number of observed orthologs ofA. gambiae genes was tallied (numbers are from the vertical columns in Table 5). The Anopheles arm (column 3) is from Table 5. Expected orthologs and percentages for each A. gambiae chromosome arm were calculated on the basis of the length these arms (see Results). The P value determined is indicated as inverse P value whereas the correspondingP value in column 9 is taken from Table 5. Column 6 is calculated from columns 2 and 4.
Local Synteny of Adh Region Orthologs
As noted above, many genes are scrambled within the respective homologous chromosomal arms. However, a careful analysis of gene order between genes of the Adh region in the D. melanogaster 2L and their orthologs in the A. gambiae 3R gave a clear indication that a significant proportion, ∼30%, remain locally clustered with the same neighboring gene. This local synteny may also be called microsynteny, in that it apparently only entails two or three genes at a time. The patterns of both gene scrambling and microsynteny are best displayed graphically, as in Figure 1. It should be noted that because of the availability of the genome sequence, the Drosophila Adhregion genes are placed on both cytogenetic and DNA sequence scales; their orthologs in Anopheles can only be placed on the cytogenetic scale for now.
Distribution of Drosophila melanogaster genes of theAdh region and their A. gambiae orthologs on chromosome 3R. The left vertical dark blue bar represents theDrosophila Adh region, listing the names of the genes, their cytological location, and their positions on the molecular map (in megabases) according to Ashburner et al. (1999). Underlined are the genes for which corresponding Anopheles orthologs are not localized on chromosome 3R (see Table 2). The dark blue vertical boxes on the right represent the chromosome arm 3R ofAnopheles gambiae, with numbered divisions and lettered subdivisions. Clones that start with 4A3A or 4A3B are cDNAs, and the rest are STSs from chromosomal BACs. The solid lines point to the cytogenetic locations of these clones. The positions of orthologs are indicated by connecting lines between the Drosophila Adh region and Anopheles chromosome 3R. Colored lines connect A. gambiae cytogenetic gene clusters (green, 29C; pink, 31C; red, 32B; and light blue, 33C) and their respective fruit fly orthologs, also shown in the same color. The two chromosomes are not drawn to scale.
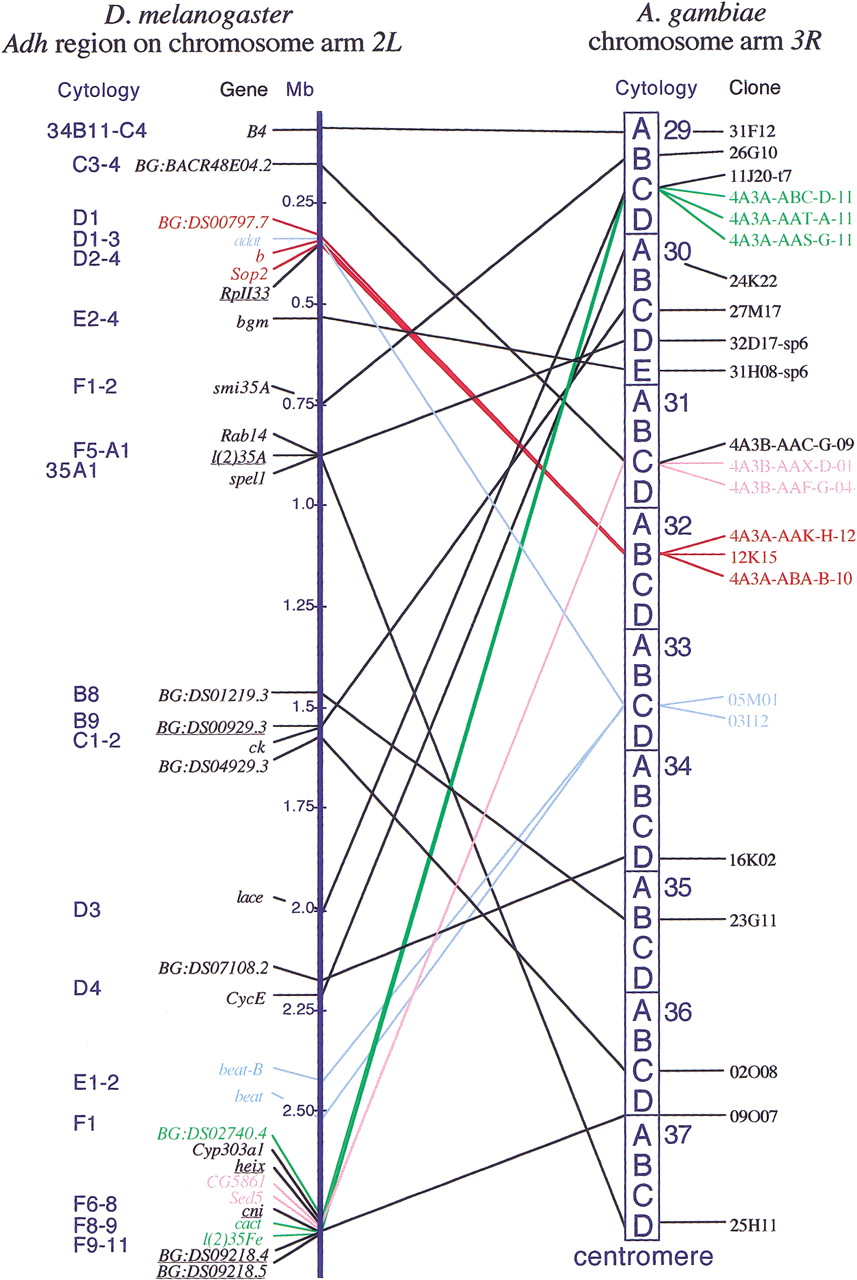
Of the 31 recognized mosquito orthologs of Adh region genes, 24 map to the Anopheles 3R chromosome, and 13 of these are found clustered in just four subdivisions, forming four cytogenetic clusters that are at least partially microsyntenic. In contrast, the remaining 11 mosquito orthologs are scattered individually amongst the remaining 33 chromosomal subdivisions of the Anopheles 3Rchromosome.
The two distal-most mosquito cytogenetic clusters, on divisions 29C and 31C, are both derived from a tight cluster of 27 Drosophilagenes that are located within ∼150 kb at cytogenetic location 35F6-11 (Ashburner et al. 1999). Of these 27 genes, 10 have known mosquito orthologs, and seven of these map to the mosquito chromosome arm3R; five are microsyntenic. The latter include two adjacent genes (the CG5861 and Sed5 orthologs) that map to the 31C cytogenetic cluster. The 29C cytogenetic cluster includes two adjacent genes (the cact and l(2)35Fe orthologs) plus one outlier (the BG:DS02740.4 ortholog). Each of these clusters additionally encompasses one ortholog of a distantAdh region gene (BG:BACR48E04.2 and lace, respectively).
Similarly, the mosquito 32B cytogenetic cluster includes threeAnopheles orthologs of genes BG:DS00797.7,b, and Sop2 that in Drosophila are part of an 16-gene cluster located within ∼65 kb at 34D1-4 (Ashburner et al. 1999). Two orthologs of other genes from the same cluster,adat and RpII33, are known in the mosquito but do not map at 32B; the orthologs of the 11 remaining genes in the 34D1-4Drosophila cluster are as yet unknown.
Finally, the fourth mosquito cytogenetic cluster at 33C includes the orthologs of adat from the Drosophila 34D1-4 region (see above) plus two genes, beat-B and beat from theDrosophila 35E1-F1 region. In Drosophila, the latter two genes are paralogs with the same exon-intron structure and show 53% identity at the amino acid level. They are separated by ∼100 kb, a region that encompasses three other genes, BG:DS07486.2,beat-C (also a paralog of beat-B and beat), and Bic-C (Ashburner et al. 1999); the orthologs of these three genes are not yet known in the mosquito. Interestingly, the orthologs of beat-B and beat are from the STSs at the two ends of the same mosquito BAC clone (03I12), and thus are also separated by ∼120 kb. It would be interesting to sequence this clone and thus discover whether the mosquito orthologs of theBG:DS07486.2, beat-C, and Bic-C genes are also located in this interval.
DISCUSSION
The analysis presented here was made possible by the availability of the essentially complete sequence of the D. melanogastergenome (Adams et al. 2000) and is a clear example of comparative genomic research. It illustrates how full genomic information from a model species can help provide considerable insight into the genomic structure of even a rather distantly related and little-studied orphan organism, when combined with bioinformatics analysis of partial sequence information and physical mapping of clones representing ESTs and STSs. It should be recalled that the fruit fly and the mosquito are estimated to have diverged ∼250 Myr (Yeates and Wiegmann, 1999). The study addresses three main questions.
The question of sequence divergence between orthologous genes ofDrosophila and Anopheles relates to our ability to detect such genes. We have used rather stringent similarity criteria to accept genes as orthologs, and thus we expect that our reported collection of orthologs includes few if any false positives and excludes some widely divergent orthologs. Consistent with these expectations, the STS resource of BAC ends represents ∼7% of the estimated euchromatic DNA of A. gambiae and yielded 26 (5.5%) orthologs of the 475 Drosophila genes present at the tip of the X and the Adh region of Drosophila. The EST resource includes 2380 cDNA clone clusters, but it is difficult to say how many actual genes are represented, because of the possibility of undetected overlaps. The EST resource yielded 24 of the orthologs or 5.1% of the genes in the Adh region and the tip of theX in Drosophila. Accepting the orthology of all genes shown in Tables 1, 2, and 4, we note that the detected orthologous exons show a range of 26% to 97% sequence identity at the amino acid level, with an average of 61.6% identity. If we consider only the most similar available exons, the orthologous genes have 31% to 97% local sequence identity, or 65.4% on average. This indicates that in most future cases, it should be possible to recognize orthologous genes in the two species using our criteria or to clone them by sequence homology.
The second question concerns the gross homology of chromosomes between the fruit fly and the mosquito. It is striking that both species have two major metacentric autosomes as well as an apparently telocentricX chromosome in the euchromatic polytene genome (five chromosomal arms in total). Only the very minor chromosome4 (∼1% of the genome in Drosophila) is absent from Anopheles. Taken together, our data show unequivocally that the five A. gambiae chromosome arms can be assigned a distinct homolog in the chromosomal complement of the fruit fly, and vice versa.
From Table 6, it can be seen that in different chromosomal arms, between 27 and 59% of the genes have undergone interchromosomal translocation to nonhomologous arms since the last common ancestor ofD. melanogaster and A. gambiae. The extent to which translocations occur varies for different arms (Table 6) and also apparently for different chromosomal regions. Comparison between Tables3 and 5 indicates that translocations have occurred more frequently for genes that are now at the X-tip of Drosophila than for the X as a whole; whereas translocations have occurred less frequently for the Adh region than for that arm as a whole. Overall, using Muller's definition of the chromosomal elements of Drosophila (Muller 1940), the A. gambiaechromosome arms X, 2R, 2L, 3R, and3L are homologous to the Drosophila elementsA, E, D, B, and C, respectively. Interestingly, in both species the arrangement of paired elements is the same (A, B + C, D + E). The A. gambiaechromosomes 2 and 3 are homologous to the D. melanogaster chromosomes 3 and 2 respectively.
A dense collection of DNA markers from Aedes aegypti(restriction fragment length polymorphisms) was used by Severson et al. (1994) to evaluate genetic diversity and synteny among aedine mosquitoes and A. gambiae; however, synteny withDrosophila was not examined. In a valuable earlier study,Matthews and Munsterman (1994) used 29 enzyme loci to study linkage conservation amongst lower diptera (13 species of mosquitoes, not including A. gambiae) and higher diptera (D. melanogaster). In different mosquito species five to 19 loci were mapped. The investigators concluded that mosquito chromosomes are modified by paracentric inversions and interchromosomal translocations. They also noted that several amall groups of two to four enzyme loci have been conserved in linkage in both mosquitoes and the fruit fly, “although most traces of homology between the two dipteran linkages have disappeared.” In the present study, a much larger number of orthologous gene sequences, mapped by in situ hybridization to polytene chromosomes, permitted firmer conclusions: pairwise identification of homologous polytene chromosomes in A. gambiae and D. melanogaster and quantification of the extent of nonhomologous arm translocations between the fruit fly and the mosquito.
The third and final issue is the distribution of genes within broadly homologous chromosomal arms, and the length of locally syntenic regions conserved between these two dipteran species. Previous studies have compared different distant Drosophila species to one another by in situ hybridization of gene-specific probes or larger genomic fragments usually derived from D. melanogaster. These studies included a cross-comparison of D. melanogaster (as a reference species) and several other species, including D. obscura,D. madeirensis, D. virilis, D. repleta,D. buzzattii, and D. hydei (Loukas and Kafatos 1988;Whiting et al. 1989; Segarra and Aguade 1992; Lozovskaya et al. 1993;Segarra et al. 1995; Nurminsky et al. 1996; Vieira et al. 1997; Ranz et al. 1999, 2000; Gonzales et al. 2000). These Drosophilaspecies were separated from D. melanogaster 25 to 60 Myr (Beverley and Wilson 1984; Russo et al. 1995). The homologous chromosome arms are usually easily identified by their gene content, but the relative order and distances of the genes are considerably reshuffled in the different species. Observed sizes of chromosomal fragments conserved between species range from 20 to 600 kb (Ranz et al. 1999, 2000; Gonzales et al. 2000), although one cannot exclude undetected small rearrangements within the larger fragments. Calculations that take into consideration the number of inversion breakpoints in several selected genomic regions and the divergence time between species indicate that the frequency of breakpoints occurring in the genus Drosophila may be as high as 0.05 to 0.08 per megabase of sequence per million years (Ranz et al. 2000). The lower estimate of this frequency would imply that in the genome of A. gambiae, calculated to have a size of ∼260 Mb, we may expect microsyntenic regions conserved relative to Drosophila to have an average DNA length of 50 to 80 kb of DNA. This is in striking contrast to the frequency of breakpoints computed for a mouse-human comparison (divergence time ∼112 Myr; Kumar and Hedges 1998), which is about two orders of magnitude lower (Ranz et al. 2000). The sizes of conserved segments in these two species are estimated to be 24 kb to 90.5 Mb in length, averaging 15.6 Mb (Lander et al. 2001). We have detected microsyntenic blocks of two to three genes each by cytological co-localization of these genes in the same Anopheles polytene chromosome lettered subdivision. It must be stressed that this evidence neither establishes nor excludes that the genes are located next to each other in the genome. As yet, we have a DNA distance estimate for only one microsyntenic pair, beat and beat-B: 100 kb in Drosophila and a BAC length (∼120 kb average) inAnopheles. However, our evidence strongly argues that locally syntenic regions between the mosquito and the fruit fly are not long.
Microsynteny between Anopheles and Drosophila was also detected by Romans et al. (1999), who isolated and characterized a 4.2-kb genomic fragment containing the Anopheles Bb,TU37B2, and Dox-A2 genes. These are orthologs of theDrosophila genes CG10655, CG10470, andDox-A2, respectively, all located within a 4.5-kb genome region in the fruit fly (Adams et al. 2000). Analysis of the molecular organization of two mosquito chromosomal regions indicated the occurrence of several rearrangements that changed both the position and orientation of Bb and TU37B2 in comparison to theirDrosophila orthologs. We have confirmed these results and found that the syntenic area does not extend much beyond the genes mentioned (data not shown).
Taking these results together, the degree of observed microsynteny between Drosophila and Anopheles is not high and may be even lower than predicted. The degree of microsynteny is an important parameter for future efforts to use the D. melanogaster gene order to identify mosquito orthologs definitively, leading to functional hypotheses and to assays of these proposed functions in the genetically tractable fruit fly. Firm elucidation of the degree of microsynteny will be one of the major benefits expected from full sequencing of the A. gambiaegenome, which is expected to begin shortly.
METHODS
Source of Sequence Data
Amino acid sequences of the genes in divisions 1–3 of chromosomeX of D. melanogaster can be obtained by anonymous FTP fromftp://ftp.ebi.ac.uk/pub/databases/edgp/misc/ashburner/EG_genes.991229.pep.fa.gz(Benos et al. 2000, 2001), whereas amino acid sequences of the genes identified in the Adh region are found in http://www.fruitfly.org/sequences/aa_Adh.dros (Ashburner et al. 1999). Amino acid sequences of all genes identified through the whole genome sequence (release 1.0) are available athttp://www.fruitfly.org/sequence/dlMfasta.html (Adams et al. 2000). ForA. gambiae, nucleotide sequences of ESTs from immune-competent cell line cDNA libraries (Dimopoulos et al. 2000) and STSs from the BAC genomic library (C. Roth and F.C. Collins, pers. comm.), as well as other mosquito sequences with known cytological location, can beBLAST-searched at AnoDB, the Anopheles database (http://konops.anodb.gr/cgi-bin/blast2.pl).
Computational Methods and Analysis of Results
For similarity searches, a locally installed WU-BLAST, version 2.0a, suite of programs (Altschul et al. 1990; W. Gish, unpubl.) was used. D. melanogaster amino acid sequences of genes from selected regions were compared to A. gambiae STS and EST databases using TBLASTN with standard default parameters. STS and EST sequences showing similarity with a high score of >40, a probability P(N) of < e−10, and a percentage of identical amino acids >30, were selected and checked as best bidirectional hits after confirming the hit usingBLASTX with standard default parameters against a database of 14,080 amino acid sequences of known and predictedDrosophila genes (release 1.0,http://www.fruitfly.org/sequence/dlMfasta.html#rel1; Adams et al. 2000). Only STSs and ESTs that passed these criteria were selected, and their alignments were further verified using the available exon-intron structure of the corresponding D. melanogaster genes, as shown in the National Center for Biotechnology Information (NCBI) version of the D. melanogaster database (http://www.ncbi.nlm.nih.gov/PMGifs/Genomes/7227.html). The sameBLASTX search criteria were also used in the reciprocal experiment, comparing A. gambiae nucleotide sequences of known cytological location to protein encoding genes in D. melanogaster genes. The names and cytological locations of D. melanogaster genes were taken from FlyBase (http://flybase.bio.indiana.edu; The FlyBase Consortium 1999); additional information and literature references on genes can also be found there.
In Situ Hybridization to A. gambiae Polytene Chromosomes
BAC and cDNA clones were hybridized to preparations of A. gambiae polytene chromosomes essentially as described in Kumar and Collins (1994). The hybridization signals were localized according to the cytological map of M. Coluzzi, A. Sabbatini, M.A. Di Deco, and V. Petrarca (unpubl., accessible at http://www.anodb.gr/AnoDB/Cytomap/).
We are indebted to Drs. Frank Collins and Charles Roth for submitting their data to public databases before publication and to Drs. Mario Coluzzi and Igor Zhimulev for their support of the participation of their laboratories in the in situ hybridization analysis of A. gambiae sequences. We would also like to acknowledge the invaluable assistance of Drs. Poulikos Prastakos and Yannis Kamarianakis in the statistical analysis. This research was supported by grants from the UNDP/World Bank/World Health Organization Special Program for Research and Training in Tropical Diseases (TDR), the INCO programme of the European Union, the National Institutes of Health, the Hellenic Secretariat General for Research and Technology, and the John D. and Catherine T. McArthur Foundation.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
Notes
[7] Corresponding author.
Notes
[8] E-MAIL [email protected]; FAX 30-81-391104.
[9] Article and publication are at http://www.genome.org/cgi/doi/10.1101/gr.196101.
REFERENCES
- ↵M. AdamsS.E. CelnikerR.A. HoltC.A. EvansJ.D. GocayneP.G. AmanatidesS.E. SchererP.W. LiR.A. HoskinsR.F. Galle(2000) The genome sequence of Drosophila melanogaster. Science 287:2185–2195.
- ↵S.F. AltschulW. GishW. MillerE.W. MyersD.J. Lipman(1990) Basic local alignment search tool. J. Mol. Biol. 215:403–410.
- ↵M. AshburnerS. MisraJ. RooteS.E. LewisR. BlazejT. DavisC. DoyleR. GalleR. GeorgeN. Harris(1999) An exploration of the sequence of a 2.9-Mb region of the genome of Drosophila melanogaster: The Adh region. Genetics 153:179–219.
- ↵C. Barillas-MuryA. CharlesworthI. GrossA. RichmanJ.A. HoffmannF.C. Kafatos(1996) Immune factor Gambif1, a new rel family member from the human malaria vector, Anopheles gambiae. EMBO J. 15:4691–4701.
- ↵C.B. BeardA.E. Crews-OyenV.K. KumarF.H. Collins(1994) A cDNA encoding an ADP/ATP carrier from the mosquito Anopheles gambiae. Insect Mol. Biol. 3:35–40.
- ↵P.V. BenosM.K. GattM. AshburnerL. MurphyD. HarrisB. BarrellC. FerrazS. VidalC. BrunJ. Demailles(2000) From sequence to chromosome: The tip of the X chromosome of D. melanogaster. Science 287:2220–2222.
- ↵P.V. BenosM.K. GattL. MurphyD. HarrisBarrellC. FerrazS. VidalC. BrunJ. DemaillesE. Cadieu(2001) From first base: The sequence of the tip of the X chromosome of D. melanogaster, a comparison of two sequencing strategies. Genome Res. 11:710–730.
- ↵N.J. BesanskyJ.A. BedellM.Q. BenedictO. MukabayireD. HilfikerF.H. Collins(1995) Cloning and characterization of the white gene from Anopheles gambiae. Insect Mol. Biol. 4:217–231.
- ↵S.M. BeverleyA.C. Wilson(1984) Molecular evolution in Drosophila and higher diptera, II: A time scale for fly evolution. J. Mol. Evol. 21:1–13.
- ↵F. CatterucciaT. NolanT.G. LoukerisC. BlassC. SavakisF.C. KafatosA. Crisanti(2000) Stable germline transformation of the malaria mosquito Anopheles stephensi. Nature 405:959–962.
- ↵C.J. CoatesN. JasinskieneL. MiyashiroA.A. James(1998) Mariner transposition and transformation of the yellow fever mosquito, Aedes aegypti. Proc. Natl. Acad. Sci. 95:3748–3751.
- ↵F.C. CollinsL. ZhengS.M. PaskewitzF.C. Kafatos(1997) Progress in map-based cloning of the Anopheles gambiae genes responsible for the encapsulation of malarial parasites. Ann. Trop. Med. Parasitol. 91:517–521.
- ↵M. ColuzziM.A. Di DecoV. Petrarca(1985) Chromosomal inversion intergradation and incipient speciation in Anopheles gambiae. Biol. Zool. 52:45–63.
- ↵A. della TorreG. FaviaG. MariottiM. ColuzziK.D. Mathiopoulos(1996) Physical map of the malaria vector Anopheles gambiae. Genetics 143:1307–1311.
- ↵M. DevenportC. BlassP. Eggleston(2000) Characterization of the Hox gene cluster in the malaria vector mosquito, Anopheles gambiae. Evol. Devel. 2:326–339.
- ↵G. DimopoulosA. RichmanA. della TorreF.C. KafatosC. Louis(1996a) Identification and characterization of differentially expressed cDNAs of the vector mosquito, Anopheles gambiae. Proc. Natl. Acad. Sci. 93:13066–13071.
- G. DimopoulosL.B. ZhengV. KumarA. della TorreF.C. KafatosC. Louis(1996b) Integrated genetic map of Anopheles gambiae: Use of RAPD polymorphisms for genetic, cytogenetic and STS landmarks. Genetics 143:953–960.
- ↵G. DimopoulosT.L. CasavantS. ChangT. ScheetzC. RobertsM. DonohueJ. SchultzV. BenesP. BorkW. Ansorge(2000) Anopheles gambiae pilot gene discovery project: Identification of mosquito innate immunity genes from expressed sequence tags generated from immune-competent cell lines. Proc. Natl. Acad. Sci. 97:6619–6624.
- ↵The FlyBase Consortium(1999) The FlyBase database of the Drosophila genome projects and community literature. Nucl. Acid Res. 27:85–88.
- ↵J. GonzalesE. BertranM. AshburnerA. Ruiz(2000) Molecular organisation of the Drosophila melanogaster Adh chromosomal region in D. repleta and D. buzzatii, two distantly related species of the Drosophila subgenus. Chrom. Res. 8:375–385.
- ↵A.A. JamesB.T. BeerntsenM. del CapurroC.J. CoatesJ. ColemanN. JasinskieneA.U. Krettli(1999) Controlling malaria transmission with genetically-engineered, Plasmodium-resistant mosquitoes: Milestones in a model system. Parassitologia 41:461–471.
- ↵N. JasinskieneC.J. CoatesM.Q. BenedictA.J. CornelC.S. RaffertyA.A. JamesF.H. Collins(1998) Stable transformation of the yellow fever mosquito, Aedes aegypti, with the Hermes element from the housefly. Proc. Natl. Acad. Sci. 95:3743–3747.
- ↵L. KamauW.R. MukabanaW.A. HawleyT. LehmannL.W. IrunguA.A. OragoF.H. Collins(1999) Analysis of genetic variability in Anopheles arabiensis and Anopheles gambiae using microsatellite loci. Insect Mol. Biol. 8:287–297.
- ↵V. KumarF.H. Collins(1994) A technique for nucleic acid in situ hybridization to polytene chromosomes of mosquitoes in the Anopheles gambiae complex. Insect Mol. Biol. 3:41–47.
- ↵S. KumarS.B. Hedges(1998) A molecular timescale for vertebrate evolution. Nature 392:917–920.
- ↵E.S. LanderL.M. LintonB. BirrenC. NusbaumM.C. ZodyJ. BaldwinK. DevonK. DewarM. DoyleW. FitzHugh(2001) Initial sequencing and analysis of human genome. Nature 409:860–921.
- ↵G.C. LanzaroY.T. ToureJ. CarnahanL. ZhengG. DoloS. TraoreV. PetrarcaK.D. VernickC.E. Taylor(1998) Complexities in the genetic structure of Anopheles gambiae populations in West Africa as revealed by microsatellite DNA analysis. Proc. Natl. Acad. Sci. 95:14260–14265.
- ↵C. Louis(1999) Malaria entomology: Can genomics help? Parassitologia. 41:489–492.
- ↵M. LoukasF.C. Kafatos(1988) Chromosomal locations of actin genes are conserved between the melanogaster and obscura groups of Drosophila. Genetica 76:33–41.
- ↵E.I. LozovskayaD.A. PetrovD.L. Hartl(1993) A combined molecular and cytogenetic approach to Drosophila evolution using large-fragment DNA cloning. Chromosoma 102:253–266.
- ↵T.C. MatthewsL.E. Munsterman(1994) Chromosomal repatterning and linkage group conservation in mosquito karyotypic evolution. Evolution 48:146–154.
- ↵H.J. Muller(1940) Bearings of the Drosophila work on systematics. in The new systematics, ed J. Huxley(Clarendon Press, Oxford), pp 185–268.
- ↵D.I. NurminskyE.N. MoriyamaE.R. LozovskayaD.L. Hartl(1996) Molecular phylogeny and genome evolution in the Drosophila virilis species group: Duplications of the Alcohol Dehydrogenase gene. Mol. Biol. Evol. 13:132–149.
- ↵S.J. O'BrienM. Menotti-RaymondW.J. MurphyW.G. NashJ. WienbergR. StanyonN.G. CopelandN. JenkinsJ.E. WomackJ.A. Marshal Graves(1999) The promise of comparative genomic in mammals. Science 286:458–481.
- ↵H. RansonB. JensenX. WangL. PrapanthadaraJ. HemingwayF.H. Collins(2000) Genetic mapping of two loci affecting DDT resistance in the malaria vector Anopheles gambiae. Insect Mol. Biol. 9:499–507.
- ↵J.M. RanzM. CaceresA. Ruiz(1999) Comparative mapping of cosmid and gene clones from a 1.6 Mb chromosomal region of Drosophila melanogaster in three species of the distantly related subgenus Drosophila. Chromosoma 108:32–43.
- ↵J.M. RanzF. CasalisA. Ruiz(2000) How malleable is the eucaryotic genome? Extreme rate of chromosomal rearrangement in the genus Drosophila. Genome Res. 11:230–239.
- ↵A.M. RichmanP. BuletC. HetruC. Barillas-MuryJ.A. HoffmannF.C. Kafatos(1996) Inducible immune factors of the vector mosquito Anopheles gambiae: Biochemical purification of a defensin antibacterial peptide and molecular cloning of preprodefensin cDNA. Insect Mol. Biol. 5:203–210.
- ↵P. RomansW.C. BlackR.K. SakaiR.W. Gwadz(1999) Linkage of gene causing malaria refractoriness to the Diphenol oxidase-2 on chromosome 3 of Anopheles gambiae. Am. J. Trop. Med. Hyg. 60:22–29.
- ↵C.A.M. RussoN. TakezakiM. Nei(1995) Molecular phylogeny and divergence time of Drosophilid species. Mol. Biol. Evol. 12:391–404.
- ↵C. SegarraM. Aguade(1992) Molecular orgenization of the X chromosome in different species of obscura group of Drosophila. Genetics 130:513–521.
- ↵C. SegarraE.R. LozovskayaG. RiboM. AguadeD.L. Hartl(1995) P1 clones from Drosophila melanogaster as markers to study the chromosomal evolution of Muller's A element in two species of obscura group of Drosophila. Chromosoma 104:129–136.
- ↵D.W. SeversonA. MoriY. ZhangB.M. Christensen(1994) The suitability of restriction fragment length polymorphism markers for evaluating genetic diversity among and synteny between mosquito species. Am. J. Trop. Med. Hyg. 50:425–432.
- ↵R.E. Sinden(1999) Plasmodium differentiation in the mosquito. Parassitologia 41:139–148.
- ↵N. TerrynP. RouzeM. Van Montagu(1999) Plant genomics. FEBS Lett. 452:3–6.
- ↵Y.T. TouréV. PetrarcaS.F. TraoréA. CoulibalyH.M. MaigaO. SankaréM. SowM.A. Di DecoM. Coluzzi(1998) The distribution and inversion polymorphism of chromosomally recognized taxa of the Anopheles gambiae complex in Mali, West Africa. Parassitologia 40:477–511.
- ↵J. VieiraC.P. VieiraD.L. HartlE.R. Lozovskaya(1997) A framework physical map of Drosophila virilis based on P1 clones: Applications in genome evolution. Chromosoma 106:99–107.
- ↵J. VizioliP. BuletM. CharletC. LowenbergerC. BlassH.-M. MuellerG. DimopoulosJ. HoffmannF.C. KafatosA. Richman(2000) Cloning and analysis of a cecropin gene from the malaria vector mosquito, Anopheles gambiae. Insect Mol. Biol. 9:75–84.
- ↵D. VlachouG. LycettI. Sidén-KiamosC. BlassR.E. SindenC. Louis(2001) Anopheles gambiae laminin interacts with the P25 surface protein of Plasmodium berghei ookinetes. Mol. Biochem. Parasit. 112:229–237.
- ↵R. WangF.C. KafatosL.B. Zheng(1999) Microsatellite markers and genotyping procedures for Anopheles gambiae. Parasitol. Today 15:33–37.
- ↵R. WangL. ZhengY.T. TouréF.C. KafatosT. Dandekar(2001) When genetic distance matters: Measuring genetic differentiation at microsatellite loci in whole genome scans of recent and incipient mosquito species. Proc. Natl. Acad. Sci. 98:10769–10774.
- ↵J.H. WhitingM.D. PlileyJ.L. FarmerD.E. Jeffery(1989) In situ hybridization analysis of chromosomal homologies in Drosophila melanogaster and Drosophila virilis. Genetics 122:99–109.
- ↵D.K. YeatesB.M. Wiegmann(1999) Congruence and controversy: Towards the higher-level phylogeny of Diptera. Ann. Rev. Entomol. 44:397–428.
- ↵L.B. ZhengR.D. SaundersD. FortiniA. della TorreM. ColuzziD.M. GloverF.C. Kafatos(1991) Low-resolution genome map of the malaria mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. 88:11187–11191.
- ↵L.B. ZhengF.H. CollinsV. KumarF.C. Kafatos(1993) A detailed genetic map for the X chromosome of the malaria vector, Anopheles gambiae. Science 261:605–608.
- ↵L.B. ZhengM.Q. BenedictAJ. CornelF.H. CollinsF.C. Kafatos(1996) An integrated genetic map of the African human malaria vector mosquito, Anopheles gambiae. Genetics 143:941–952.
- ↵L. ZhengA.J. CornellR. WangH. ErfleH. VossW. AnsorgeF.C. KafatosF.C. Collins(1997) Quantitative trait loci for refractoriness of Anopheles gambiae to Plasmodium cynomolgi B. Science 276:425–428.