
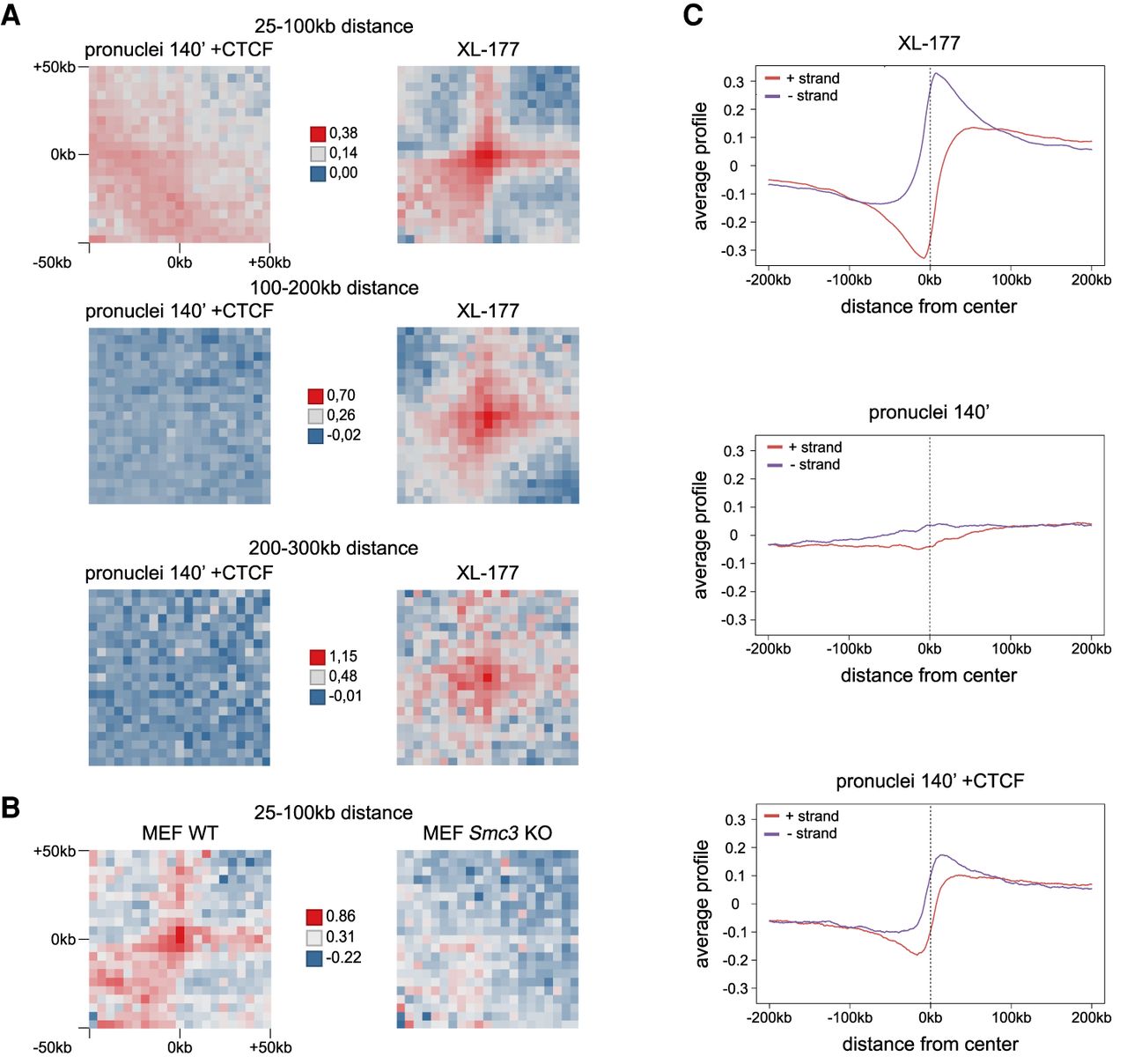
Addition of recombinant CTCF to Xenopus pronuclei fails to induce typical corner peak or TAD formation but generates weak contacts between Smc3 sites at very small distances. (A) Log2 of observed/control of aggregate contact frequencies between consecutive, directly adjacent Smc3 binding sites with a distance between the binding sites as indicated and 5-kb bin size, for demembranated frog sperm in Xenopus egg extract after 140 min (pronuclei 140′), pronuclei 140′ with addition of 50 nM CTCF protein, and frog XL-177 cells. To calculate control values, Smc3 binding sites were shifted by 10 Mb. (B) Log2 of observed/control of aggregate contact frequencies for mouse WT MEF cells and Smc3 KO MEF cells between consecutive, directly adjacent Smc3 binding sites in WT MEF cells with a distance between the binding sites of 25–100 kb and 5-kb bin size. Hi-C data by Banigan et al. (2023); ChIP-seq data from this study. To calculate control values, Smc3 binding sites were shifted by 10 Mb. (C) Average profiles of the directionality index for XL-177 cells, pronuclei 140′, and pronuclei 140′ with addition of 50 nM CTCF protein over unambiguously oriented CTCF binding sites detected in pronuclei 140′ +CTCF.








