
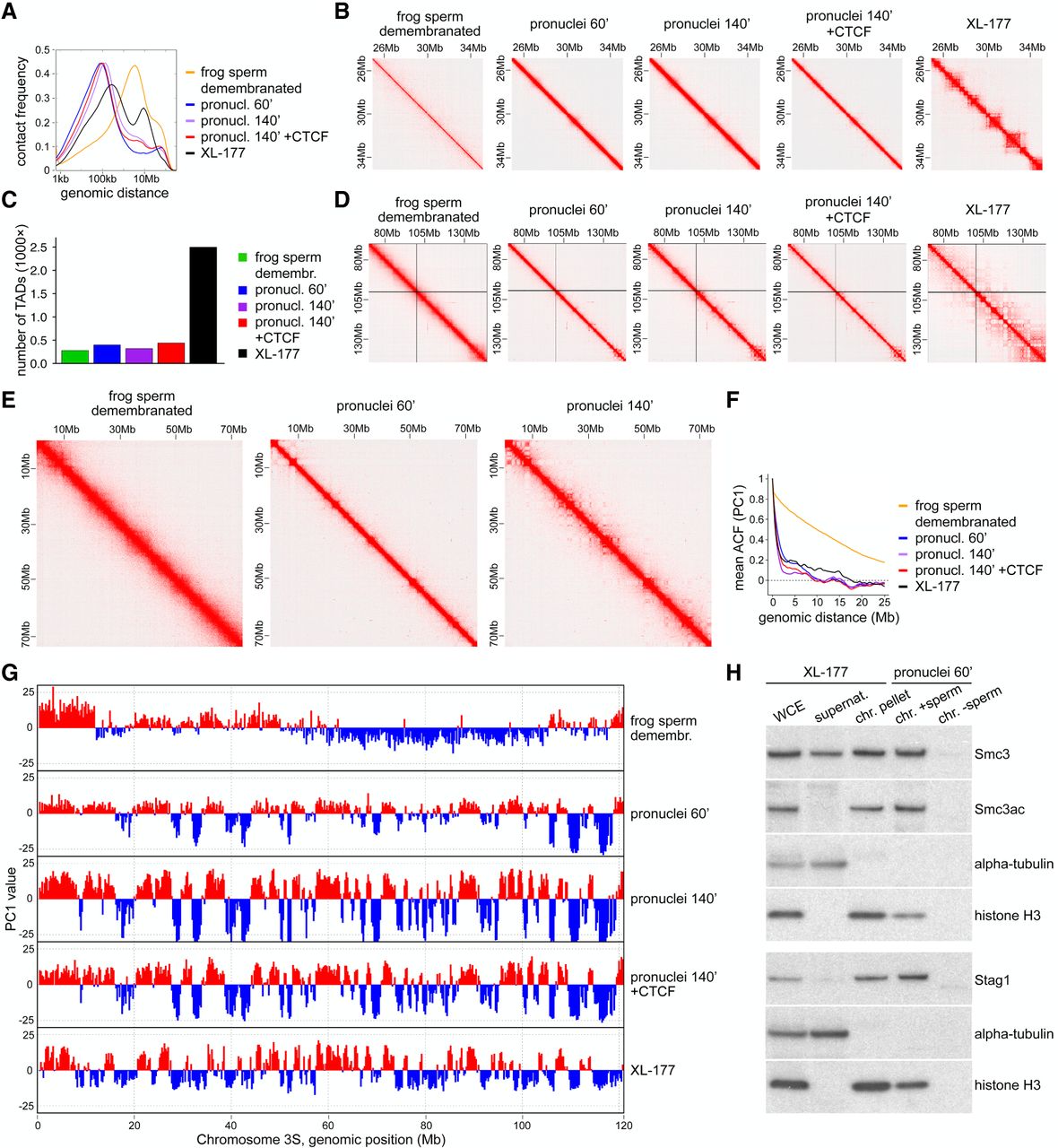
Demembranated frog sperm is remodeled upon incubation in egg extract. (A) Contact frequency as a function of genomic distance for demembranated frog sperm, demembranated frog sperm in Xenopus egg extract after 60 min (pronuclei 60′) or 140′ (pronuclei 140′), pronuclei 140′ with addition of 50 nM CTCF protein, and frog XL-177 cells. (B) Normalized Hi-C matrices for the same samples as in A in the region of Chr1S:25–35 Mb at 25-kb resolution. (C) Number of TADs called for the same samples as in A. (D) Normalized Hi-C matrices for the same samples as in A in the region of Chr3L:70–150 Mb at 100-kb resolution. (E) Normalized Hi-C matrices for three of the samples in A in the region of Chr1S:0–75 Mb at 100-kb resolution. (F) The autocorrelation of the PC1 value as a function of genomic distance, for the same samples as in A. (G) Compartment tracks from principal component analysis for the same samples as in A. (H) Immunoblotting of chromatin isolated from pronuclei 60′, with or without sperm, in comparison with chromatin fractioning (whole cell extract, supernatant, chromatin pellet) of XL-177 cells.








