
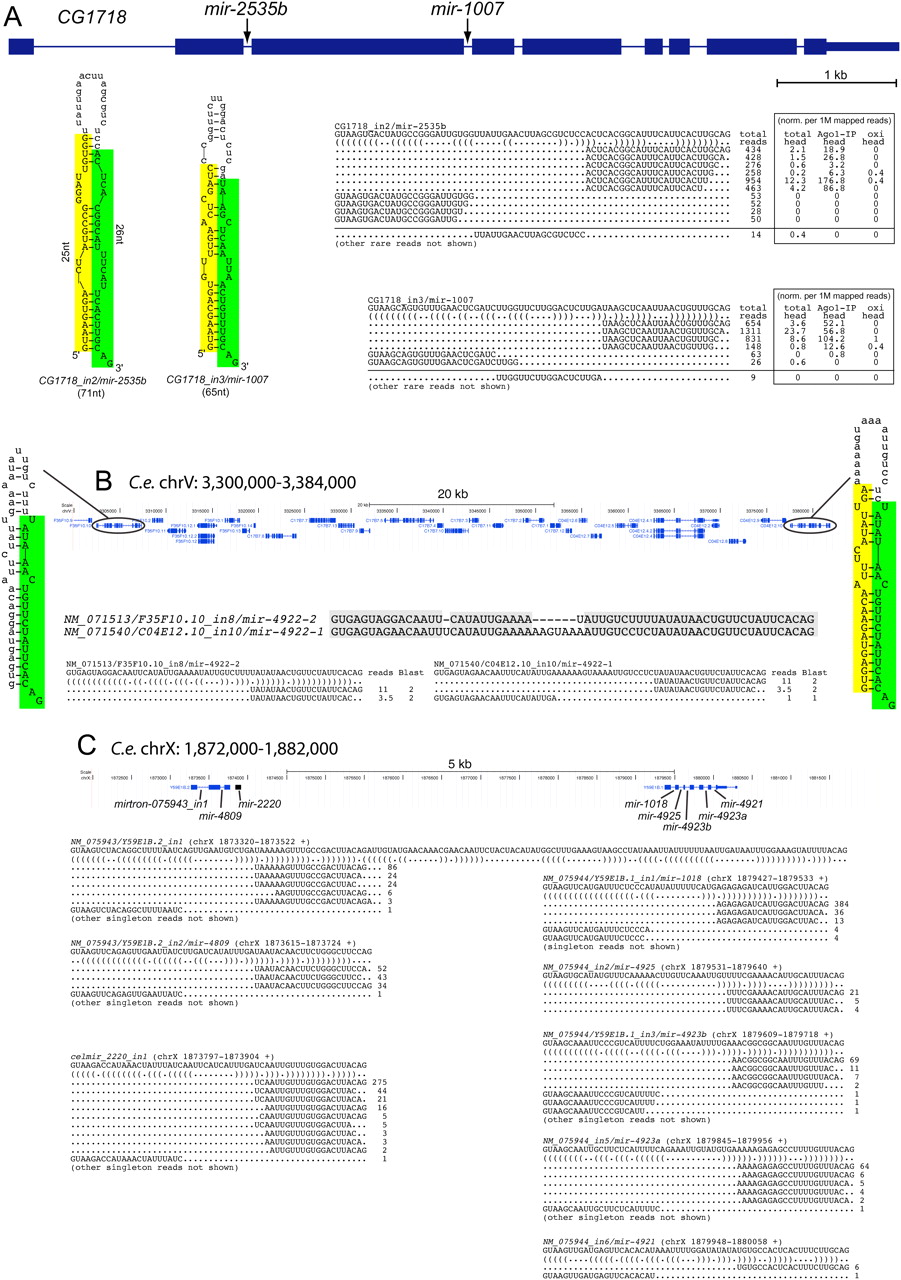
Clustered mirtrons in the D. melanogaster and C. elegans genomes. (A) Drosophila CG1718 generates mirtrons from both its second and third introns; CG1718_in2 was newly identified in this study. Curiously, while the hairpin structure of CG1718_in2 is seemingly suboptimal compared with the previously identified mir-1007, mature miRNAs accumulate to relatively similar levels from these mirtrons. Analysis of head libraries published by Ghildiyal et al. (2010) provided evidence that these mirtrons are expressed in the head and generate RNAs that populate AGO1, but not AGO2 complexes; this study used oxidation (oxi) of input samples to enrich for 2'O-methylated RNAs in mature AGO2 complexes. To permit comparison between the total and IP levels, these read numbers were normalized per million mapped reads in each library. Rarer reads were not shown, except for the informative cloned terminal loops that report on endogenous Dicer-1 processing; the full read patterns are available at http://cbio.mskcc.org/leslielab/mirtrons. (B) NM_071513 and NM_071540 are related genes that reside ∼70 kb apart on C. elegans chromosome V. Each gene bears a mirtron whose 3p arm is identical; thus, small RNA reads from this arm map to both mirtrons. We normalized the read numbers to assign half to each locus. On the basis of unique star arms, we can definitively annotate the expression of NM_071540. However, given that the hairpin of NM_071513 has only small symmetric loops, we infer that its processing should be equivalent, if not more efficient, to its paralog. (C) A supercluster of mirtron genes on C. elegans chromosome X. This <8-kb region was previously annotated to contain mir-1018 and mir-2220, of which mir-1018 was previously noted to be a mirtron (Ruby et al. 2007a). Although mir-2220 was earlier annotated as a canonical miRNA (Kato et al. 2009), we infer that it is similarly a mirtron, as its cloned RNAs begin and end with effective splice junctions. Here, we identify six additional mirtrons in this genomic region. Of these, NM_075943_in1 might appear to be a tailed mirtron based on the annotated splice junction; however, that its abundant 3p reads end with CAG suggests that it may be the product of alternative splicing, as seen for the Drosophila mirtron CG17560. Note that in all gene alignments only a subset of informative singleton reads, typically belonging to mirtron star species are shown.








