
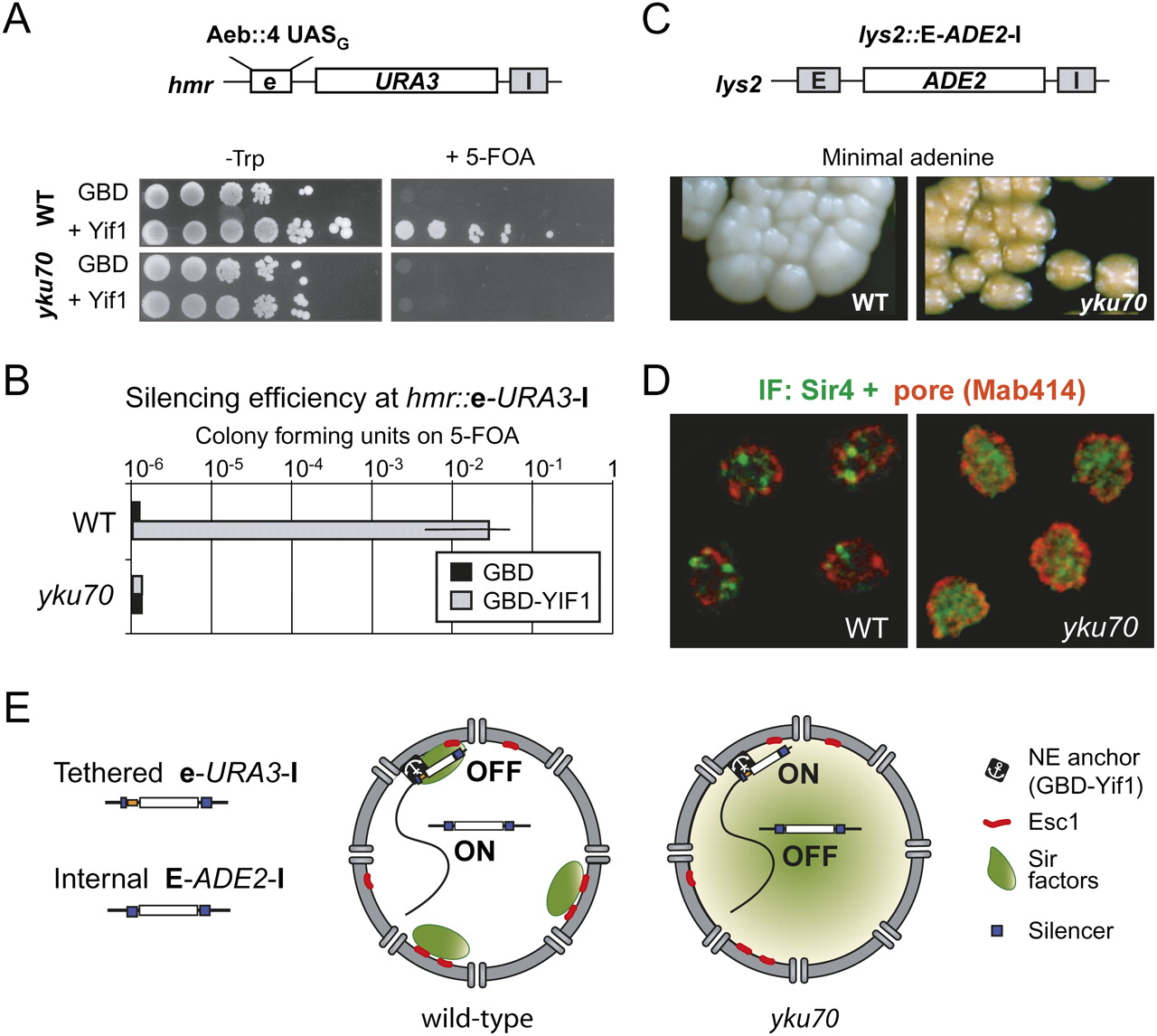
SIR distribution affects both perinuclear silencing and ectopic silencing. (A) Targeted silencing is monitored in strains carrying URA3 at HMR adjacent to a modified E silencer (hmr∷e-URA3-I) in which the E and B elements are replaced by four UASG sites (Chien et al. 1993). Serial 10-fold dilution series of GA-2511 (WT) and its yku70 deletion derivative (GA-2585) expressing different GBD fusions from a high copy TRP1 plasmid are grown on SD-trp with or without 5-FOA, to monitor URA3 repression. (B) Bar graph representing the mean and standard deviation for ratios of growth in the presence or absence of 5-FOA, for four independent transformants. Indicated cell backgrounds all carried the hmr∷e-URA3-I reporter. (C) Ectopic silencing is monitored in the same strains bearing an ADE2 gene bracketed by the E and I HML silencers integrated at LYS2, in the middle of chromosome 2 (lys2∷E-ADE2-I). Wild-type colonies are white due to full ADE2 expression, while yku70 colonies are pink, indicative of partial ADE2 silencing. (D) Immunolocalization of Sir4 and nuclear pores (MAb414) in the strains GA-1998 (WT) and GA-1997 (yku70), expressing GBD–Yif1. Bar = 1 μm. (E) Model for the effect of SIR protein distribution on silencing. The lefthand nucleus reflects a wild-type yeast cell in which SIR proteins are sequestered at telomeric foci (green areas), such that internal reporters are not silent unless they are anchored at the NE by a transmembrane protein (e.g., GBD–Yif1; Andrulis et al. 1998). In the righthand nucleus, the SIR proteins are distributed throughout the nucleus due to ablation of Ku. In this strain, tethering the crippled silencer to the nuclear periphery does not promote silencing. On the other hand, local recruitment of SIR proteins by a strong silencer can promote repression at an ectopic position.








