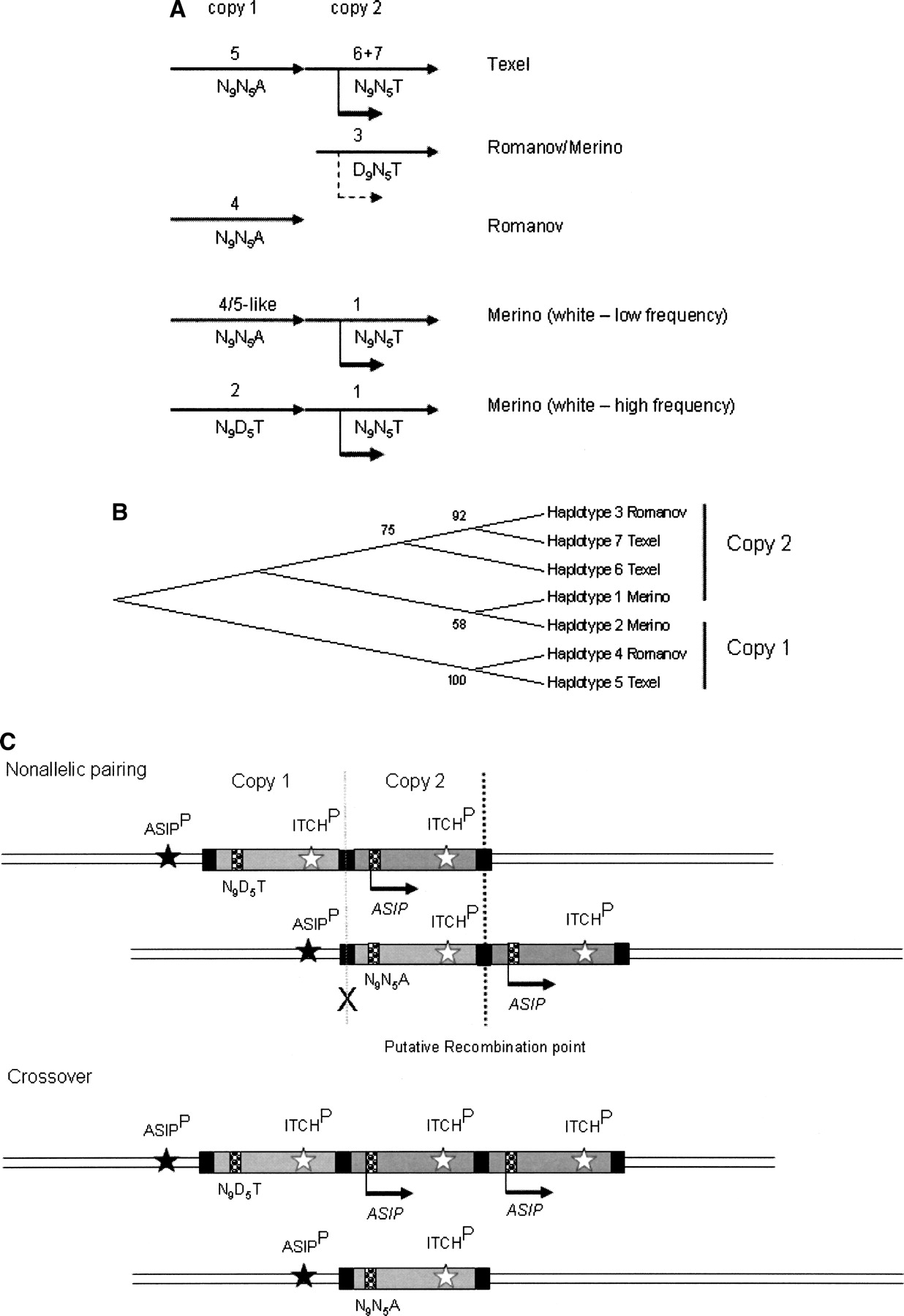
A model for the organization and evolution of the ovine ASIP locus in dominant white and recessive black domestic sheep. The proposed positions (A) and clustering (B) of copy 1 (inactive) and copy 2 (active) haplotypes identified from Texel, Romanov, and Merino sheep are shown. Solid arrows indicate high levels of expression of haplotypes at copy 2. A dashed arrow indicates expression of a haplotype 3-like D9 allele detected in Merino skin. The two haplotypes identified in the Romanov BAC library may not be derived from a duplicated ASIP-ITCH region as pure Romanov sheep have a recessive black-like phenotype. However, haplotype 4 clusters with haplotype 5 in copy 1 of the Texel and haplotype 3 clusters with haplotype 6 and 7 from the Texel and haplotype 1 from the Merino (B). This clustering suggests that the Romanov haplotypes are derived from the two different copies of the putative original gene duplication. The low frequency presence of a haplotype 4/5-like copy 1 region in the Merino sheep population is deduced from the identification of N9N5A genotypes in the black Merinos and is consistent with the organization of the region in the Texel sheep CHORI-243 library BACs. (C) Schematic showing resolution of nonallelic pairing between duplicated ASIP-ITCH copies with crossover products showing reciprocal deletion and triplication. The 190-kb segments of copy 1 and copy 2 of a white Merino are shown as light and dark gray boxes, respectively. Black boxes indicate the similar SINE sequence regions at the junction, 5′ and 3′ breakpoints. In this example, the ASIP genotype shown represents a white animal heterozygous at copy 1 for the nonfunctional alleles (N9D5T/N9N5A) and homozygous at copy 2 for the functional alleles (N9N5T/N9N5T). Arrows below the ASIP genes indicate the direction of transcription driven by the ITCH promoter (ITCHP). Nonallelic pairing and crossover between the junction and 3′ breakpoints (dark dashed line) would result in creation of a single copy nonfunctional (N9N5A) allele, as shown. The resulting single ASIP copy is not expressed as the ancestral ASIP promoter (ASIPP) is silent. A crossover point (gray cross) that would result in the positioning of a nonfunctional (N9N5A) allele under the regulation of the duplicated ITCH promoter is unlikely to occur as expression of “A” alleles was not detected in Merino sheep. Variable positioning of functional haplotypes at copy 1 and 2 combined with nonallelic pairing and crossover explains the different single copy alleles identified in black Merinos. Mutations and gene conversion events could also contribute to the diversity at the locus.







