
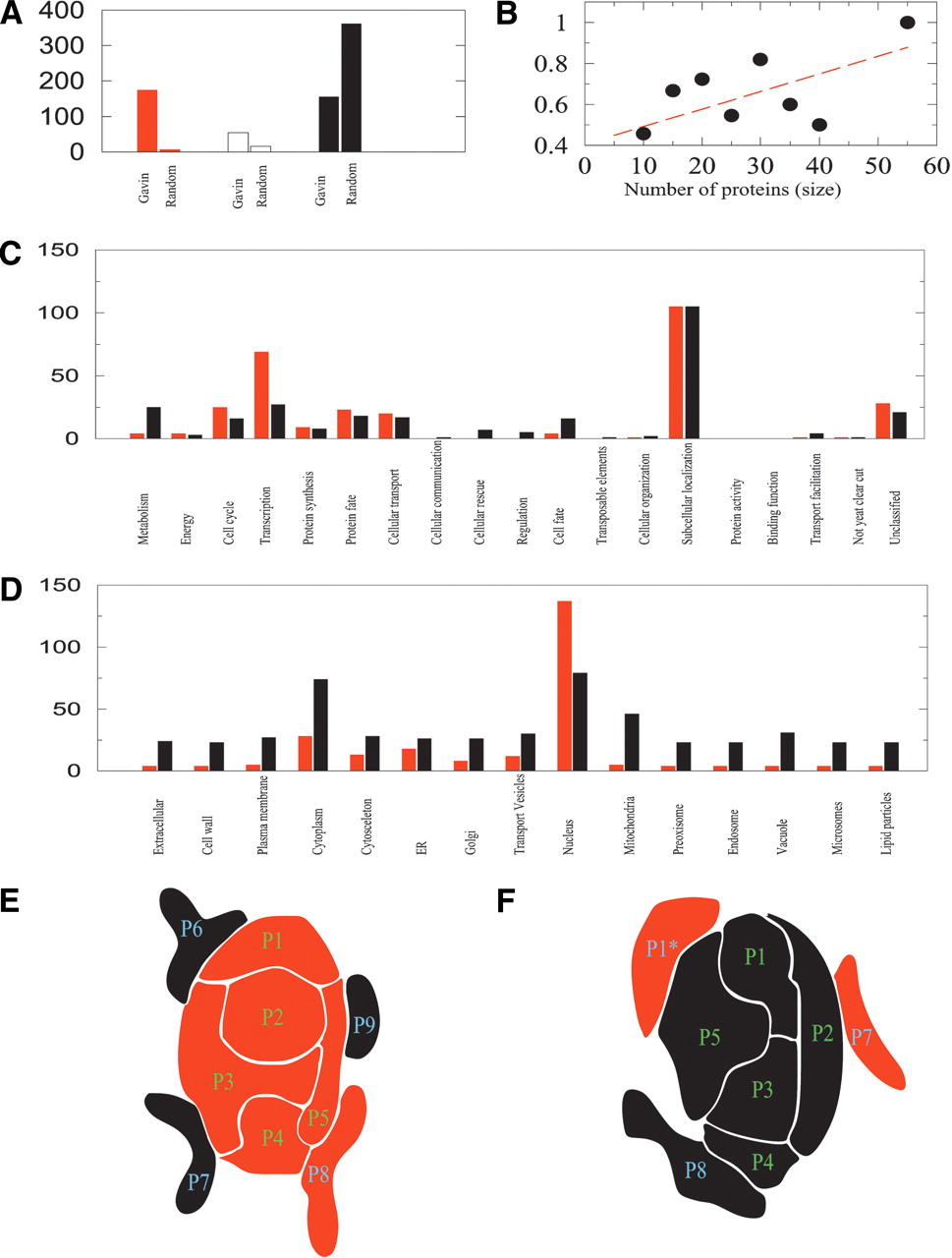
Characterization of the protein complex ensemble and schematic illustration of the internal organization of protein complexes. (A) The number of complexes in the Gavin et al. (2002) data set that are found to be essential (red), nonessential (black), and of unknown (white) deletion phenotype. Next to each column we show the number of corresponding complexes if the proteins were randomly distributed in the various complexes, indicating the highly nonrandom character of the complex composition and essentiality. For this, each protein subunit of the known Gavin et al. (2002) complexes is replaced with proteins randomly selected from the yeast proteome. (B) The size dependence of essentiality in protein complexes. The plot shows the fraction of essential subunits within the complex (vertical axis) as a function of the number of protein subunits in the complexes (horizontal axis). (C) The predicted functional classification of the complexes identified by Gavin et al. (2002), showing separately the number of essential and nonessential complexes found in each functional class. (D) The predicted cellular localization of the identified essential and nonessential complexes. A full list of predictions for each complex is shown in the Supplemental material. (E) We find that 43% of the protein complexes possess a core comprised of highly coexpressed proteins, that are all essential and belong to the same functional class, indicating that they represent the functional building blocks of the complex. Such core is shown schematically as tightly locked P1-P5 proteins. Mass spectroscopic methods inevitably identify other proteins as well with those complexes. Yet, we find that these halo proteins (P6-P9) show a small coexpression pattern with the core, and are both phenotypically and functionally mixed, indicating that they likely represent proteins that display only temporal or spurious attachment to the complex. (F) Approximately 46% of the complexes have a core composed of predominantly nonessential proteins (P1-P5), surrounded again by a halo of proteins with mixed essentiality and functional classification (P6-P8). These complexes likely are not essential for cell growth; therefore, all core proteins are uniformly nonessential. The few essential proteins found predominantly in the halo of such nonessential complexes often simultaneously take part in the core of other essential complexes, explaining the origin of their essentiality. For example, the P1 protein, which is part of the core of the essential complex shown in E, could also attach to the surface of the nonessential complex shown in F. Therefore, the essentiality of P1* is derived not from its role in complex F, but from its role in the essential complex E.








