
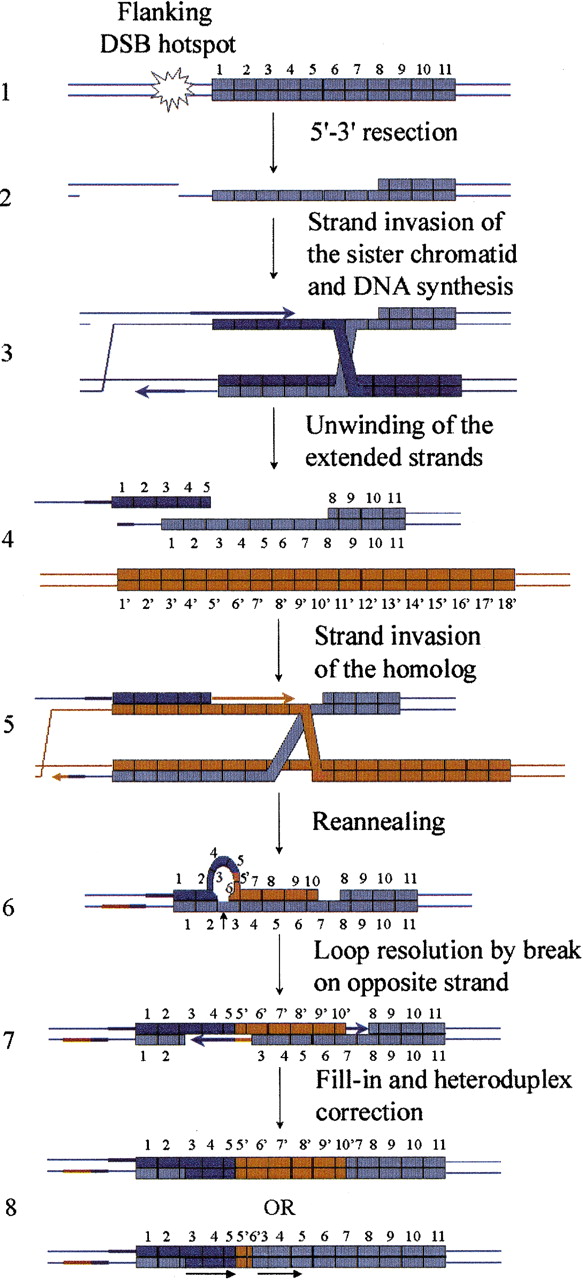
Revised model for meiotic mutation events demonstrating the formation of interallelic events with duplications flanking the converted motifs. In order to explain the observed duplication flanking meiotic interallelic events, the model initially proposed by Buard and Vergnaud (1994) and subsequently adopted by others (Bois and Jeffreys 1999) invoked DSBs initiated within the array by staggered single-strand breaks separated by 80 nucleotides or more. This would require a strong associated helicase activity and does not fit with the view now provided by the yeast work (Debrauwère et al. 1999). Alternatively, the presented model adapted from Debrauwère et al. (1999) shows how an almost blunt DSB, produced in a flanking DSB hotspot outside the minisatellite (step 1), can produce interallelic exchanges with a duplication flanking the converted motifs, as well as most, if not all, minisatellite rearrangements observed in man or yeast. After 5′-3′ resection (step 2), the repair is initiated by invading the sister chromatid and priming DNA synthesis on one or both (as suggested here, step 3) strands. After DNA synthesis, the newly synthesized strands independently unwind (step 4) and are free to engage in other DNA–DNA interactions (here, both strands are shown invading the homolog, step 5). Eventually, the newly synthesized strands reanneal together with properly aligned flanking sequences. A loop may form on one (as shown here, step 6) or both (Debrauwère et al. 1999) strands. This loop can be converted into the corrected allele (Debrauwère 2000) via a single strand cut on the opposing DNA strand (step 7), or removed. Depending on which strand is used to correct the heteroduplex, a direct duplication of repeats flanking the converted patch is produced (step 8, bottom). All models proposed so far predict the existence of patches of heteroduplex intermediates produced by the reannealing of similar, but different, minisatellite motifs (here, step 6). This last prediction was successfully tested inDebrauwère et al. (1999). An interesting aspect of this model is that the lower strand may extend in the flanking sequence at steps 2 and 4. This will produce a heteroduplex region in the flanking sequence (steps 6 and 7; left unrepaired here in step 8) which, once repaired, may introduce a conversion patch in the final product flanking sequence.








