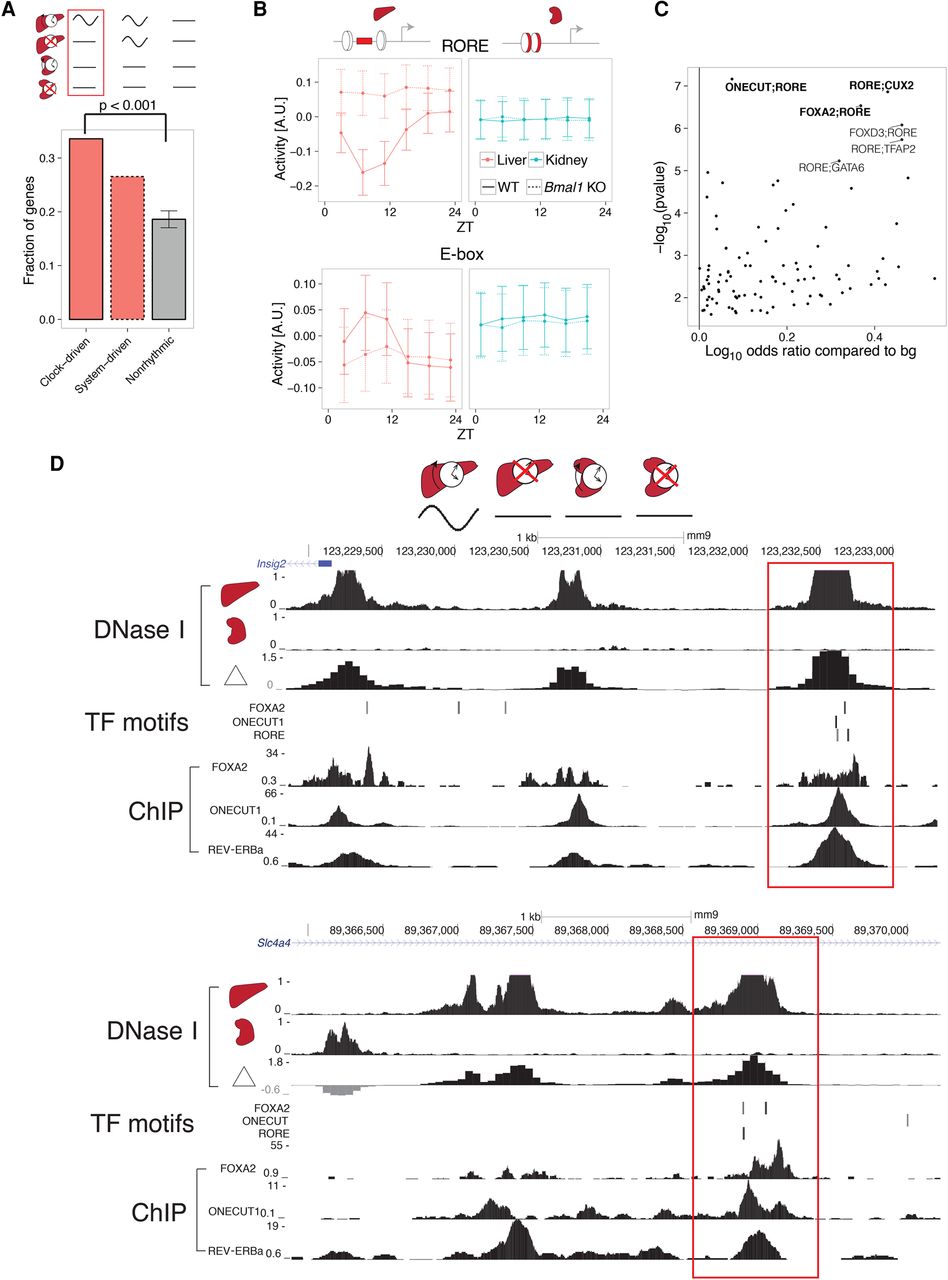
Colocalized binding of clock- and liver-specific TFs underlies liver-specific mRNA rhythms. (A) The fraction of genes containing liver-specific DNase I hypersensitive sites (DHSs) in the clock-driven liver-specific module is higher compared with both nonrhythmic and system-driven liver-specific modules. Error bars and P-values calculated from 10,000 bootstrap iterations. (B) Predicted temporal activities of RORE (top) and E-box (bottom) TF motifs located within liver-specific DHSs. Error bars show standard deviation of the estimated activities. (C) Co-occurrence of RORE with all other TFs in the SwissRegulon database (Pachkov et al. 2007) (189 TF motifs). Positive log10 odds ratios (ORs) represent pairs of motifs enriched in the clock-driven liver-specific module compared to the flat module. P-values for the motif pairs were calculated from χ2 tests applied to three-way contingency tables (Myšičková et al. 2012). Selected pairs are in bold. (D) DNase I hypersensitivity in liver, kidney, and the corresponding differential signal (in log2 fold change) near two representative genes (top: Insig2; bottom: Slc4a4). RORE, ONECUT1, and FOXA2 TF binding motifs (posterior probability > 0.5, MotEvo) co-occur at liver-specific DHSs (red boxes). Predicted TF binding sites correspond to experimentally observed TF binding in publicly available ChIP-exo data sets for REV-ERBa, ONECUT1, and FOXA2 (bottom).