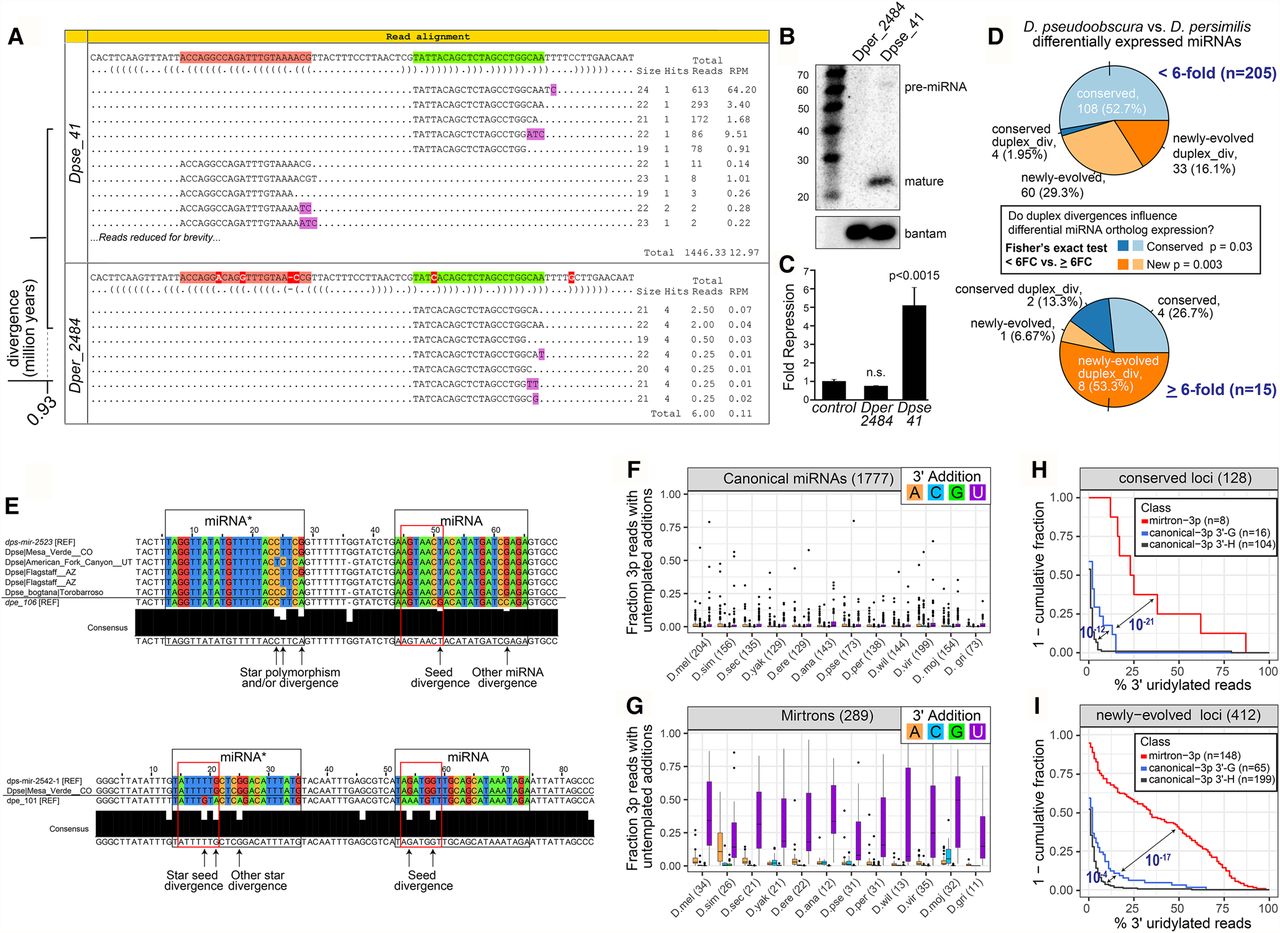
Multiple distinct cis-molecular signatures associated with miRNA flux. (A–C) Duplex alterations affect miRNA maturation and function. (A) The Dpse_41/Dper_2484 ortholog pair, with only a few duplex divergences, exhibits divergent expression between very closely related species. Functional assays confirm differential biogenesis of Dpse_41/Dper_2484 expression constructs by Northern blotting (B) and differential activity by luciferase sensor assay (C). (D) Transcriptome comparison of miRNAs differentially expressed between sister species Dpse and Dper. In general, significantly more duplex divergent miRNAs are differentially expressed miRNAs for both newly evolved and conserved miRNAs. (E) Evolution of seed regions of testes-restricted, clustered (TRC) miRNAs. Shown are examples of one-to-one orthologs of TRC miRNAs between Dpse and Dper, including available Dpse population data. Highlighted are examples of seed divergence between expressed TRC miRNA orthologs between these closely related species, consistent with adaptive evolutionary behavior. (F–I) Impact of terminal uridylation system on evolutionary suppression of mirtrons and behavior of canonical miRNAs. (F,G) Compared to canonical miRNAs (F), mirtrons (G) in every Drosophilid species acquire much higher rates of terminal untemplated uridylation (purple) on the 3′ ends of their 3p species, compared to any other nucleotide modifications. (H,I) 3′ uridylation of canonical miRNAs is sensitive to terminal hairpin nucleotide. In these graphs, miRNA loci are divided by biogenesis type (canonical versus splicing-derived), by terminal nucleotide (3′-G versus 3′-A/U/C, i.e., “3′-H”), and by evolutionary age. Analysis of deeply conserved miRNA loci (H) and recently evolved loci (I) shows that canonical miRNA hairpins that end in G acquire higher levels of 3′ uridylation than do other canonical miRNA hairpins. P-values computed from a two-tailed Wilcoxon rank-sum test.