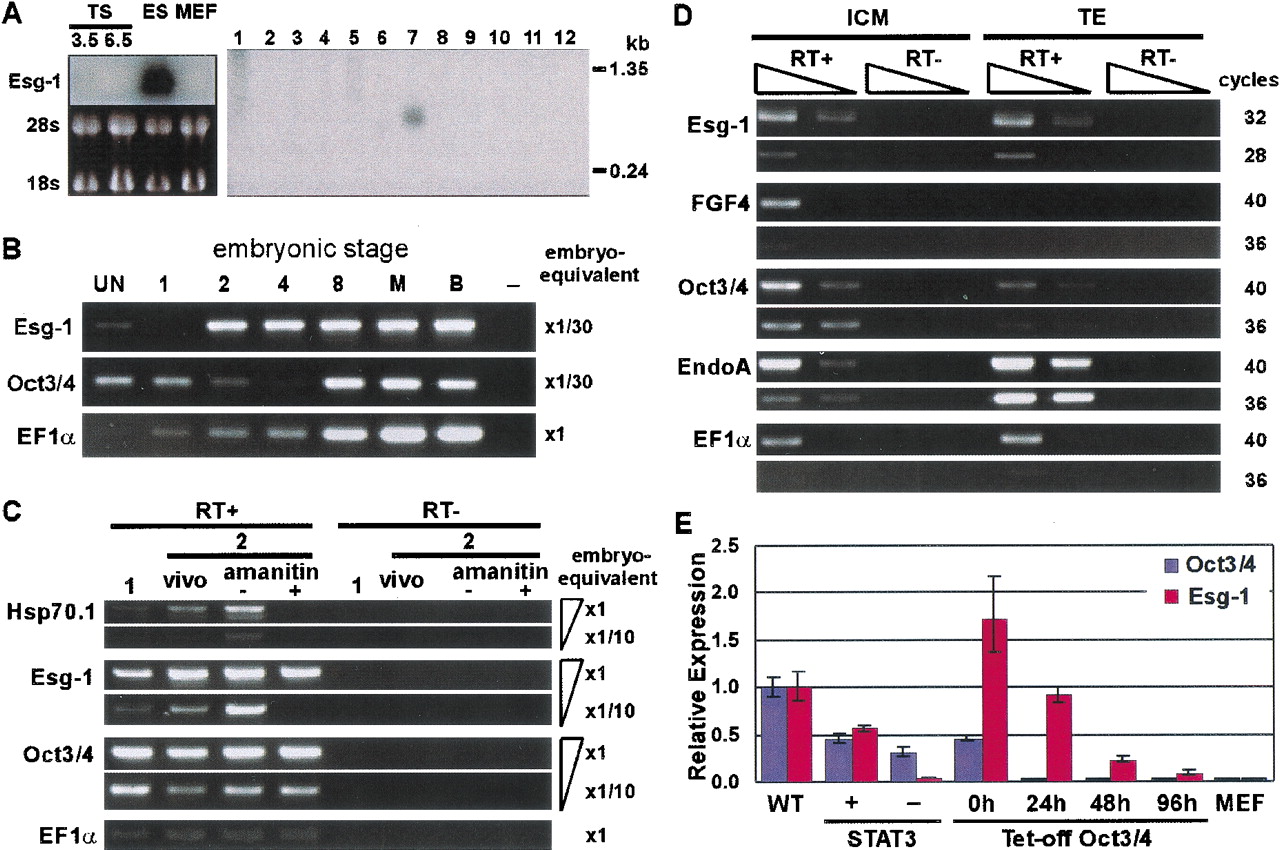
(A) Northern hybridization of Esg-1. (Left) Total RNA from ES, TS3.5, TS6.5, and MEF cells were blotted onto a nylon membrane. The image of ribosomal RNA is shown as a loading control. (Right) poly(A)+ RNA from multiple cell-lines were blotted onto a nylon membrane (Clontech). Cell lines used for this blot are (1) neuroblastoma NB41A3; (2) mastocytoma P815; (3) lymphoma P388D1; (4) lymphocytic leukemia L1210; (5) lymphoma R1.1; (6) hepatoma Hepa1–6; (7) embryonic carcinoma P19; (8) subcutaneous connective tissue-type L-M; (9) fibroblast M-MSVBALB/3T3; (10) fibroblast k-BALB; (11) Abelson murine leukemia virus-induced tumor RAW264.7; (12) lymphoid tumor PU5–1.8. (B–D) Semi-quantitative RT-PCR analysis in preimplantation embryos. Primer pair for EF1α was used as a loading control. (B) Expression patterns ofEsg-1 and Oct3/4 were examined in unfertilized eggs (UN; 278 eggs), fertilized eggs (1; 248 zygotes), and preimplantation embryos at stages of 2-, 4-, and 8-cells (289, 312, and 181 embryos, respectively), morula (M; 144 embryos), and blastocyst (B; 175 embryos). It was confirmed that RT negative controls of each sample gave no PCR products. Only RT negative control of blastocyst (−) is shown. Each cDNA was first adjusted to one egg or one embryo-equivalent and then serially diluted as indicated to the right. (C) Early zygotic transcription-dependent expression ofEsg-1 was confirmed by α-amanitin treatment of fertilized eggs. Fertilized eggs with a polar body were collected at 27 h post-hCG injection and either uncultured (1; 313 zygotes), or cultured for 18 h in M16 with (+; 207 embryos) or without (−; 230 embryos) 100 μg/mL α-amanitin. Two-cell stage embryos (vivo; 226 embryos) were also collected directly from oviducts in parallel. Serially-diluted cDNAs were used as indicated to the right. Embryos were pooled from four series of experiments. Hsp70.1 was used as a positive control. On average, 75% and 67% of the fertilized eggs developed to two-cell stage embryos in the absence and presence of α-amanitin, respectively. (D) Expression level of Esg-1 in the ICM and TE was investigated by direct isolation of the ICM and TE by immunosurgery and FITC-ConA labeling followed by trypsinization, respectively. A combination of serial dilution of cDNAs and different PCR-cycles were indicated. Fgf4 and Oct3/4 were known to be expressed in the ICM, whereas EndoA is a marker of the TE (Brulet and Jacob 1982). (E) Real-time RT-PCR analysis ofEsg-1 and Oct3/4 in manipulated ES cells. Two manipulated ES cell-lines either expressing active Stat3 conditionally, or suppressing Oct3/4 conditionally were used as sources of RNA. MEF cells were used as a negative control. The expression level ofOct3/4 was also measured as a marker of pluripotency. The expression levels of Esg-1 and Oct3/4 were normalized to those of EF1α, whose constant expression levels over different conditions were verified (data not shown). The levels ofEsg-1 and Oct3/4 in the manipulated cell lines were compared with those of nonmanipulated ES cells grown under standard conditions.