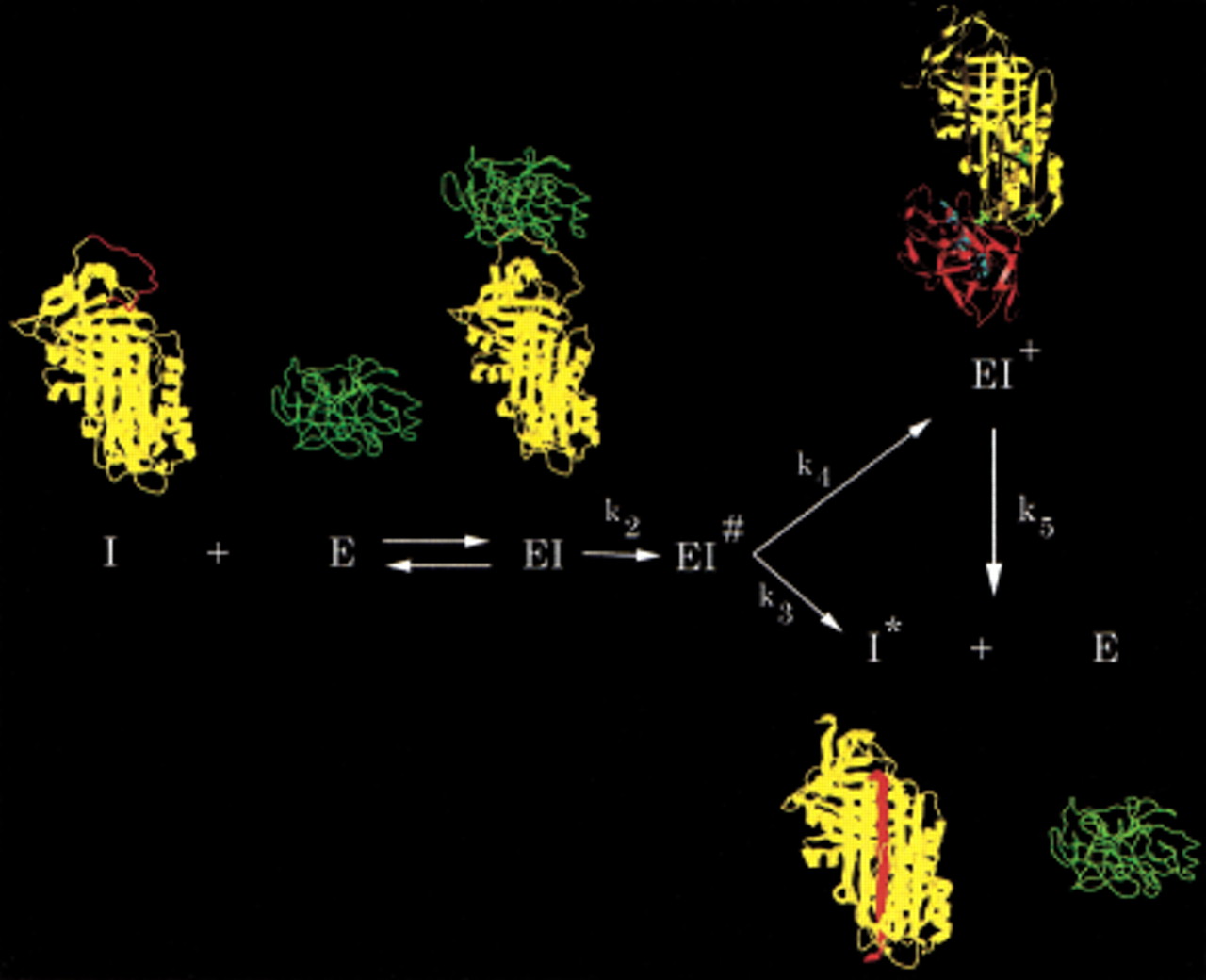
Branched pathway mechanism of serpin (I) inhibition of proteinase (E), illustrated using α1PI as the serpin and trypsin as the proteinase. Serpin and proteinase bind to form an initial noncovalent Michaelis-like complex (EI) in which the proteinase docks with the exposed reactive center loop (red) with no distortion of the proteinase or the body of the serpin (Peterson et al. 2000). Normal substrate-like cleavage occurs with rate constant k2 to form the acyl enzyme intermediate EI#. A competition ensues between the inhibitory pathway (rate constant k4) and the substrate pathway (rate constant k3). Completion of the substrate reaction by hydrolysis of the acyl enzyme intermediate results in release of cleaved serpin (I*) and free proteinase. The inhibition pathway involves insertion of the cleaved reactive center loop into β-sheet A of the serpin and concomitant dragging of the covalently bound proteinase from the top to the bottom of the serpin to give the kinetically trapped covalent serpin–proteinase complex (EI+; Stratikos and Gettins 1999;Huntington et al. 2000). In the covalent complex, the proteinase (shown in red in the EI+ structure) is distorted through compression against the bottom of the serpin and, hence, rendered catalytically incompetent (Huntington et al. 2000). Because the catalytic competence of the proteinase has been severely compromised by the distortions within the active site, completion of the hydrolysis of the acyl intermediate from the trapped complex (rate constant k5) is extremely slow. In vivo complexes are cleared before appreciable dissociation has time to occur.