Abstract
The debate on modern human origins has often focused on the relationship between genes and fossils. Although more and more genetic evidence has been accumulating in favor of a recent African origin for modern humans, it has been assumed by many that the fossil evidence remains ambiguous. On the contrary, it has been clear for some time that the fossil evidence does not support the multiregional model: Fossils and archeology indicate a pattern of multiple dispersals from and beyond Africa, against which the genetic data can be compared. The continuing value of paleobiology is in complementing genetic information by revealing the context of human evolution: locating the dispersals and extinctions of populations in time and space, correlating these events with the environmental forces that shaped them, and providing an increasingly detailed understanding of the morphology and technology of early humans.
Molecular biology has revolutionized the study of human evolution. The importance of fossils as the primary source of information about our past has been steadily undermined as it has become possible to infer detailed aspects of recent human history from the distribution and frequency of genes found around the world today. To some extent, a fusion of paleontological and genetic approaches came about last year with the extraction and sequencing of ancient DNA from an extinct hominid, the Neanderthal type specimen (Krings et al. 1997), but this is likely to remain a rarity. Ancient DNA, however, has been able to confirm that humans and Neanderthals belonged to different populations over the last quarter of a million years and that Neanderthals did become effectively extinct.
These results fit in with an increasingly consistent interpretation of human evolutionary genetics. Compared with chimpanzees and other apes, the human population is relatively lacking in genetic diversity (Ruvolo et al. 1993); such genetic variation as does exist occurs primarily within populations rather than between (Relethford and Harpending 1994); African populations are more diverse genetically than those found anywhere else in the world (Vigilant et al. 1991; Cavalli-Sforza et al. 1994; Watson et al. 1997); and for the most part, non-African patterns of genetic variation can be treated as a subset of African ones. The chronological and demographic context of the processes of diversification has been strongly disputed, but several genetic systems indicate that living populations derive from a relatively small population (effective population size between 5000 and 50,000 individuals) (Rogers and Harpending 1992; Harpending et al. 1993; Nei and Takahata 1993; Harpending 1994). That size represents a bottleneck in the hominid lineage dating back no more than 200,000 years (Cann et al. 1987; Stoneking et al. 1992), with evidence of demographic expansion occurring in the last 70,000 years (Rogers 1995).
This emerging consensus has been interpreted in evolutionary terms as evidence for a recent African origin (i.e., ∼200,000 years ago) for modern humans, followed by a dispersal out of Africa, with little or no interbreeding with other populations of hominid. Many anomalies and further details remain to be sorted out. The geographical clades of the mtDNA have been questioned (Maddison 1991; Maddison et al. 1992), and it has been suggested that much of the genetic evidence is consistent with a number of different models (Templeton 1992, 1993). Furthermore, tree building and mismatch techniques, the primary means of turning genetic information into evolutionary history, have been subject to dispute (Templeton 1992, 1993). Finally, estimates of the coalescence time of human β-globin genes (∼800,000 years) have also been used to argue against a recent African origin for modern humans (Fullerton et al. 1997), but they are not, in fact, inconsistent with the recent-origin model, as the coalescence time for a neutral autosomal locus would be expected to be roughly four times that of a mitochondrial gene, and selection may also play a confounding role.
Such disputes, though, should perhaps not distract from the general pattern: Genetics supports a single recent African origin for modern humans. This consensus leads to two important questions: First, what is the relationship between this evidence and the fossil evidence that it has been claimed, supports the so-called multiregional model? And more broadly, what can paleoanthropology contribute to a field where the bulk of the resources is being put into genetics? The answer to the first question is that it has been clear for a long time that the morphological evidence does not support a multiregional view. However, this has not percolated through to the genetic community. This apparent lack of communication between the two disciplines can be used to help provide an answer to the second question: that paleobiology provides a different sort of evidence, one that focuses specifically on the context in which human genetic evolution occurred. These themes will be developed below.
Multiregional Evolution
The multiregional model of human evolution has a long history. It was first proposed by Franz Weidenreich (1943) in relation to his analyses of the Peking Man fossils but has been further developed and promoted in recent years by Milford Wolpoff and Alan Thorne (Wolpoff et al. 1984). The essence of this model is that since the origin ofHomo some two million years ago, the human lineage has evolved as a single lineage, without speciation events (Wolpoff and Caspari 1997). Within this lineage there would have been universal directional trends in characters such as brain size, but regional differences in populations would have persisted. Over time, therefore, some specific morphological traits would persist in particular regions, regardless of other global trends; the human lineage thus evolved in several regions (multiregionality). This continuity has been seen to extend to the transition from archaic to modern hominids in Africa, Asia, and Europe, the last of these being the link between Neanderthals and modern Europeans. The multiregional model is therefore largely inconsistent with the general interpretation of the genetic evidence, requiring that the effective population size of the human lineage was always large enough for gene flow to occur globally and that coalescence times for most genes would be in excess of a million years.
Paleontological Support for a Single-Origin Model
A number of paleontological observations have been consistently quoted as supporting the multiregional model and have allowed the debate on the origin of modern humans to persist. However, it is worth summarizing the results of some recent studies that bear on this issue.
Australian Populations and Homo erectus in Southeast Asia
The most often cited evidence for continuity outside Africa is that between modern Australian populations and the later specimens ofHomo erectus, the Ngangdong sample from Java (Fig. 1). Australians are supposed to share with the late-surviving H. erectus a number of cranial traits (Larnach and Macintosh 1974;Wolpoff et al. 1984). Upon superficial visual comparisons, Ngangdong and some recent Australian specimens may appear similar, but it is misleading to interpret this as part of a unique evolutionary relationship. First, the earliest known Australian fossils, those from Mungo, are quite gracile and do not show these traits (Brown 1987). The later ones, like Kow Swamp and Keilor, are very diverse, implying that some became more robust with time (Lahr 1996). Second, new dates indicate that Ngangdong may be as young as 50,000 years; if this is the case, then the supposed source population persisted unchanged long after the origins of the Australians. Third, the few characters that Australian populations do share with with Southeast Asian H. erectus are also found in the early modern human fossils of Eastern Africa (Omo Kibish) and the Levant (Skhul, Qafzeh) (Lahr 1996), and metrically, the most “primitive” of the Australian fossils (the Willandra Lakes specimen WLH50) falls closest to the African transitional or early modern specimens (Stringer 1998). These African specimens, however, also have modern proportions of the skull, unlike those found in Ngangdong that are essentially archaic and thus fall closer to Australians and represent a better ancestral model (Groves and Lahr 1994). In other words, where Australian populations look like archaic ones, it is those on the cusp of modernity in Africa that they resemble most (Fig. 1).
Fossil modern hominid skulls from Australia and Africa, compared with the H. erectus Ngangdong skull from Java. (A) Cast of Ngangdong, (B) Lake Mungo 3 (Australia); (C) Willandra Lakes 50 (WLH-50, Australia); (D) Kow Swamp (Australia): (E) Keilor (Australia); (F) Ngaloba (African transitional); (G) Omo Kibish (early modern African). Photograph credits: (A) R. Foley; (B–E) kindly supplied by Peter Brown (University of New England, Armidale, NSW, Australia); (F,G) kindly supplied by Chris Stringer (Natural History Museum, London, UK). For further information about the fossil Australian hominids, see Peter Brown’s Web page athttp://www.une.edu.au/~Arch/PalaeoIndes.html/.

Regional Continuity Traits
Central to the support for multiregional evolution is the notion that certain anatomical traits show a persistence and uniqueness within regions, regardless of the time of the specimens. These are the so-called continuity traits (Wolpoff et al. 1984). Three regional groups are usually discussed: Australian, East Asian, and European. In practice, it is the first two of these that have been the primary focus of discussion and have provided the most often-cited evidence for continuity. There has, however, only been one major study of the actual frequency, distribution, and biological significance of the regional traits (Lahr 1994, 1996). In that work, Lahr took samples from the main continental populations and looked at how many of the traits showed statistically significant specific regional distributions, that is, occurred at significantly higher levels in one region of the world rather than another. The results effectively undermine the validity of the continuity traits for showing evolutionary links between archaic and modern populations in different parts of the world. For example, of the 10 traits supposedly specific to East Asia, 2 showed no regional pattern, 6 showed higher frequencies in regions other than East Asia, and only 2 could be shown to be genuine, regionally specific characteristics of present East Asian populations. Nineteen Australian traits have been suggested, but again only six proved to be regionally specific to Australia. These results do not mean that there is no regional variation in human populations, nor do they preclude the possibility that there may be continuity in some features, but they do indicate that there is no statistical basis for the claimed regional continuity of this particular set of features.
Temporal Continuity
The implication of regional continuity is that the same traits can be found consistently in populations from one time period to the next. This is seldom, if ever, the case. The situation with Southeast AsianH. erectus and the Australians has already been described. In East Asia, the earliest modern humans are those from Upper Cave Zhoukoudian, dated to ∼30,000 years ago. Kaminga and Wright (1988)have shown that statistically these specimens differ from recent East Asians. As in Australia, the diversity of early moderns in eastern Asia refutes notions of temporal stability in regional patterns (Lahr 1996).
Morphometrics
Another way of testing hypotheses concerning the origins of modern humans is to look at overall morphological similarity and differences among Homo crania. Howells (1973, 1989) originally suggested that all modern human crania shared more in common with each other than any did with their local antecedents. Stringer (Stringer 1992a,b, 1995) has repeatedly also shown in multivariate analyses that “recent modern” and “early modern” human skulls are all distinct from nonmodern forms and do not show any trends within these regions that might indicate the evolution locally of modern humans from archaic ones. This would include such specimens as WLH50 from the Willandra Lakes in Australia, which despite being remarkably big and robust is also essentially very modern (Turbon et al. 1997). Pairwise comparison of the relevant populations also supports this view (Waddle 1994).
Cladistics
In paleontology, at least, there is a long-standing argument about the relative strengths of phenetics (simple assessment of overall similarities and differences, usually based on metrics) and cladistics (a more discriminating technique that, although generally less based on quantifiable traits, is more careful in weighting the evolutionary significance of each trait) as means of reconstructing evolutionary relationships. Phenetics is thought to be less powerful as it does not weight characters according to degree of ancestral sharing. However, the appropriateness of applying cladistics to within-species variation has been debated (Trinkaus 1990). Leiberman (1995) has carried out a cladistic analysis of the recent hominid cranial variation and has shown that the most parsimonious explanation is that which incorporates all modern human variation in a single clade, derived from a recent African source. Groves and Lahr (1994) have also shown that many of the continuity traits used in the multiregional model that supposedly link Australian and Javanese H. erectus populations are in fact shared ancestral characteristics of the Homo clade and therefore cannot provide useful phylogenetic information.
Neanderthal Admixture
Although there is growing acceptance among paleontologists that the western European Neanderthals show an extreme morphology and do not appear to have interbred with modern humans, it has been claimed that there is evidence for admixture in central Europe. A major element of that claim is the material from Krapina in former Yugoslavia, which lacks many of the more extreme Neanderthal features. However, recent redating of the material shows that it is in fact ∼130,000 years old, which appears too old for admixture with incoming humans (Rink et al. 1995), and its lack of specialization is almost certainly a result of its position early in Neanderthal evolution. Pleistocene modern Europeans cannot be shown to have special affinities with Neanderthals (Stringer et al. 1984; Howells 1989; Van Vark 1990; Van Vark and Henke 1992).
Chronology
Dating of later Pleistocene hominids has advanced considerably over recent years, and this new dating information has been a central to the debate. The key fact to emerge is that there is a chronological fault line in the appearance of modern humans. In eastern Africa, northern Africa, southern Africa, and the adjacent Levantine region, the first modern humans are over ∼100,000 years old (Stringer et al. 1989; Grun et al. 1990; Grun and Stringer 1991). In Europe, eastern Asia, and virtually everywhere else archaic hominids persist until <40,000 years ago, largely unchanged, and modern humans are not found until after these dates (Mercier et al. 1991; Hublin et al. 1996;Swisher et al. 1996). The 60,000 year or 3000 generation gap seems to suggest that African and non-African human evolution was largely decoupled until very recently.
Overall, therefore, whatever element of evidence is considered—geographical patterns, chronological distribution, or cladistic and general morphological relationships—there is little or no evidence for non-African admixture or continuity between archaic and modern populations. These paleontological results seem to be unequivocal and support fully the single African origin model of recent human evolution. Publications within paleoanthropology over the last decade have consistently failed to provide positive support for the widespread in situ evolution of modern humans. Although it is often stated that there is a conflict between genetics and fossils, there is little basis for this view, and new studies rather than older ones have tended to be entirely consistent with a single African origin for modern humans.
Genes and Fossils: Complementary Approaches
Given the apparent consistency between genetic and paleobiological data, it might be reasonable to ask what role the fossils now serve. If we can learn the same story from the genes, perhaps they should be seen as the primary source of information about recent human evolution. Paleontologists might soon join their specimens in extinction. In fact, though, the demise of the old debate between multiregional and single-origin models opens the opportunity for a much more complementary approach to the problems of human evolution. As more diverse sets of questions are asked, what each discipline can and cannot do is becoming clearer.
Paleoanthropology
The great strength of paleontology is that it can locate things in time and space and as such can provide the context in which evolutionary events occurred. As explaining evolution is largely a question of showing the links between changes in genotypes or phenotypes and the external conditions, the context is clearly very important. Explanations of why human evolution took the form it did must in the end depend on paleobiology. In contrast, paleontology cannot with any great ease talk about gene flow very precisely. Tiny amounts of genetic exchange are unlikely to be very visible. Only the broad patterns are observable. Thus, for example, when paleoanthropologists talk about replacement, they are talking about the general event, not whether very low-level genetic admixture may have occurred.
Genetics
Genetics reconstructs evolutionary events by inferring the past from the present. It is very powerful at building phylogenetic relationships within and between species and to a lesser extent at locating them in time and space. However, as currently used, genetics can say little or nothing about populations that have not persisted through to the present day and nothing about context. Furthermore, although the situation is bound to change, most evolutionary genetic studies are concerned with noncoding parts of the genome, and thus, little can be said about the functional or structural nature of present or past human biology.
The complementarity of the two disciplines is thus built on the fact that paleontology can reach the world of extinct populations that provided the context in which humans evolved and locate it in time and space, whereas genetics can provide the details of recent history. Thus far, the sole focus for genetic research has been phylogeny and timing; the mechanisms and population-level processes involved have not been addressed, largely because of the priority given to solving the “two theory problem.” With that solved, greater emphasis can now be placed on process and hence on the integration of genetic and paleontological evidence.
Multiple Events, Multiple Dispersals: Reconstructing the Context of Human Genetic Diversity
What do the fossil and archeological records reveal? BoxFigure B1. shows the climatic curve for the last half million years, on which have been highlighted a number of key events that may be relevant to the evolution of modern humans. “Single-origin” models of human evolution have tended to imply that the origin is a single event. Perhaps the first point to note about the diagram is that the fossil and archeological evidence has for some time indicated that this is not the case, and, increasingly, genetic evidence is bearing this out. Any valid model of modern human evolution must incorporate multiple events and multiple dispersals. In particular, it is clear that although anatomically modern humans may have evolved ∼150,000 years ago, modern dispersals did not occur until much later, and then there were several. Future models of human evolution based on genetics should recognize the full complexity of the events of the last quarter of a million years (Foley and Lahr 1992;Lahr and Foley 1994).
Multiple events: key points in recent human evolution. (1) The (approximate) time scale for considering the fossil evidence for the origins of H. sapiens is around half a million years. By this stage, hominid populations were well established in Africa, Europe, and Asia, and there is evidence for diversity among them. The recent estimate of the coalescence point for modern human and Neanderthal mtDNA is a little longer ago than this date, whereas the diversification of the African ancestral population leading to modern humans and that leading to European Neanderthals is likely to be more recent than this date (Krings et al. 1997). Based on fossil evidence, the likely ancestral population leading to all these half-million-year-old hominids was in Africa between 1.5 and 2.0 million years (Myr) ago. (2) The environmental context for considering later human evolution is that of repeated glacial cycles; this is shown best by the marine isotope record, which estimates the temperature of the ocean by measuring the ratio of two stable oxygen isotopes (delta 18 values on the y-axis) (Shackleton 1996). There have been several major glaciations during this period, each of which would have produced profound climatic and biogeographical changes. The last interglacial and glacial period is but one of many that have occurred, and each one is itself highly variable. During very cold periods of the glacial (glacial maximum), much of Europe would have been under glaciation, the rest open tundra and steppe. Africa would have been cold and arid, and the rain forests reduced and fragmented. During interglacials, Africa and Eurasia would have been far more forested; during periods of decreased aridity, migration of African faunas across the Sahara would have been more possible. (3) A snapshot of the hominid populations between half a million years and 300,000 years ago would have suggested the following: (1) persistence of H. erectus with a simple chopper and flake industy in Southeast Asia and possibly in parts of Europe and eastern Asia as well and (2) an Afro-European distribution of a more derived and larger brained hominid—Homo heidelbergensis—with Acheulean handaxes and associated tools; this population may also have spread into parts of eastern Asia. (4) Around 300,000 years ago a new type of stone tool technology appeared in Africa, and this was probably associated with the evolution and dispersal of a new and more adaptable hominid species, the ancestor of both humans and Neanderthals (Homo helmei). This Mode 3 technology [a Middle Stone Age/Middle Paleolithic technology based on prepared core technology, where the core or pebble that is used is prepared in advance before a flake is struck off (see Fig. 2)] persisted in the early modern humans and is probably evidence of a very much more advanced behavioral capacity. The Mode 3 or Middle Stone Age dispersals were among the first of the ones that established the modern human world (Foley and Lahr 1997). (5) The Isotope Stage 6 glacial maximum was a period of sustained and extreme cold. In Africa there would have been isolation and fragmentation of human populations, a process possibly associated with the ancestral modern human bottleneck. In Europe there may have been a virtual loss of hominid populations. It was during this period that modern human anatomy may have evolved among isolated African populations, and the first modern human fossils (the Omo Kibbish sample from Ethiopia) probably date from this period. (6) The last interglacial was a period of ∼10,000 years of warming, much of it equivalent to today’s climate. It was during this period that the fossil and archeological evidence suggest that modern human populations expanded to the south (the Klasies River Mouth sample in South Africa) and to the north, out of Africa for the first time (the Skhul and Qafzeh samples in Israel). These early modern populations may well have become locally extinct at the end of the last glaciation. One such early modern population may have dispersed through a southern route, around the Indian Ocean Rim, and into Australia and New Guinea (Lahr and Foley 1994). (7) Although not as cold as the Last Glacial Maximum, the early part of the last glaciation (∼70,000 years ago) saw the contraction of African environments and populations, their disappearance from the Middle East, and an apparent expansion of Neanderthals from Europe into areas previously occupied by modern humans in the Levant. (8) From ∼45,000 years ago, there were a number of dispersals, often associated with a new technology (Klein 1992; Foley and Lahr 1997). These were not global but a series of regional ones: the Upper Paleolithic in Europe, the Later Stone Age in sub-Saharan Africa, and one with populations using microliths in southern and eastern Asia. The subsequent 15,000 years appear to have seen recurrent expansions and contractions, and changes in population distribution. It was during this period that the last of the nonmodern populations became extinct: the Neanderthals in Europe,H. erectus in Southeast Asia, and possibly other populations in eastern Asia. (9) From 21,000 years ago to 18,000 years ago, world climates reached their coldest and driest point of the last 130,000 years, the last glacial maximum (LGM). During this period, human populations would have been under major selective pressures again. These would not have been uniform, resulting in some populations concentrating demographically in favorable enclaves, others diluting throughout vast arid landscapes, whereas still others suffered significant demographic loss. (10) The global warming out of the LGM was extremely rapid (although with a sharp reversal for a short period of time), and the succeeding demographic and geographical expansions placed human populations under new pressures for survival, out of which agriculture emerges in various parts of the world. This in turn led to increasing human population density and new dispersals, resulting in both asimilation and extinction of populations marginal to these developments.
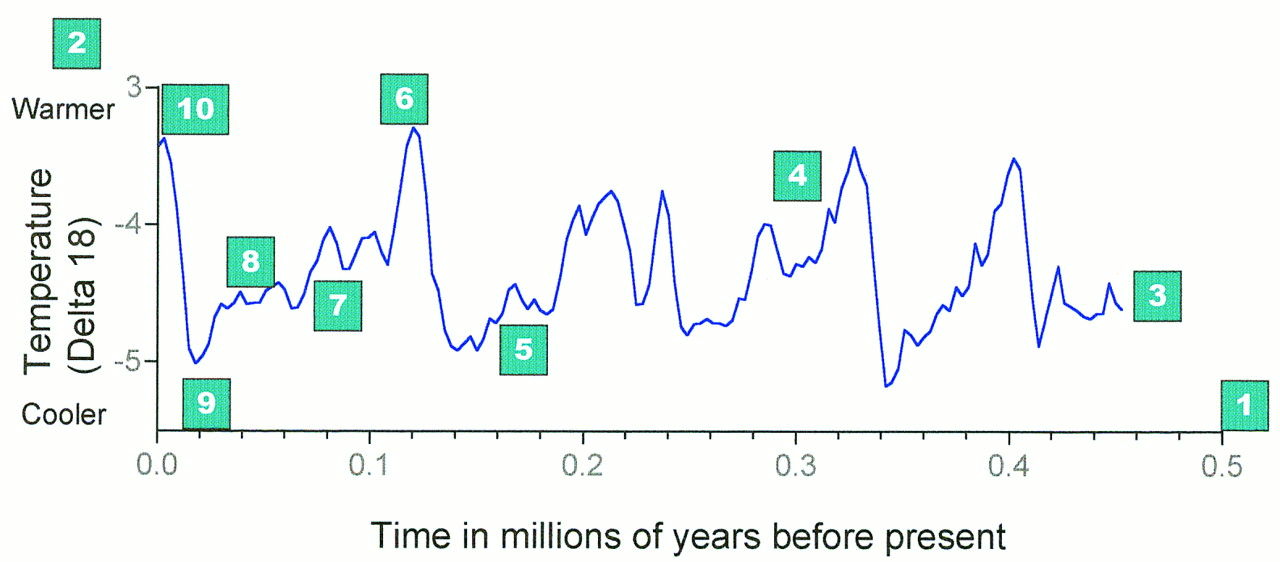
The second point is that the Neanderthals are just a small part of the story and, perhaps, distort the way we look at the past. There has been considerable discussion about whether the Neanderthals are a separate species. However, although there is more and more evidence that they did not interbreed with early European modern humans, the species concept is in practice largely irrelevant to the problems of the later Pleistocene. What the fossil record shows is a number of populations, modern and archaic, sparsely scattered across time and space (Fig.2). Many of these, modern or archaic, may well not have made it through to the present day and thus are not reflected in current genetic diversity. Given this, and the complexity of the events, caution should be maintained when looking for links between the living and the fossil populations. Essentially, we need population- not species-based models. Recent human biological history is at heart a question of microevolution.
The paleobiological context for the evolution of modern humans. This diagram shows the general pattern for the evolution of Homo.Time is on the vertical axis, and the horizontal dimension is a representation of space, with Africa (middle), Europe (left), and Asia (right). The width of each band (very roughly) represents population size. The major population divergences (speciation events) are shown, with their associated technological systems. Generally speaking, the estimated coalescence points of genes will predate the actual population divergences shown in the paleontologically based estimates here.
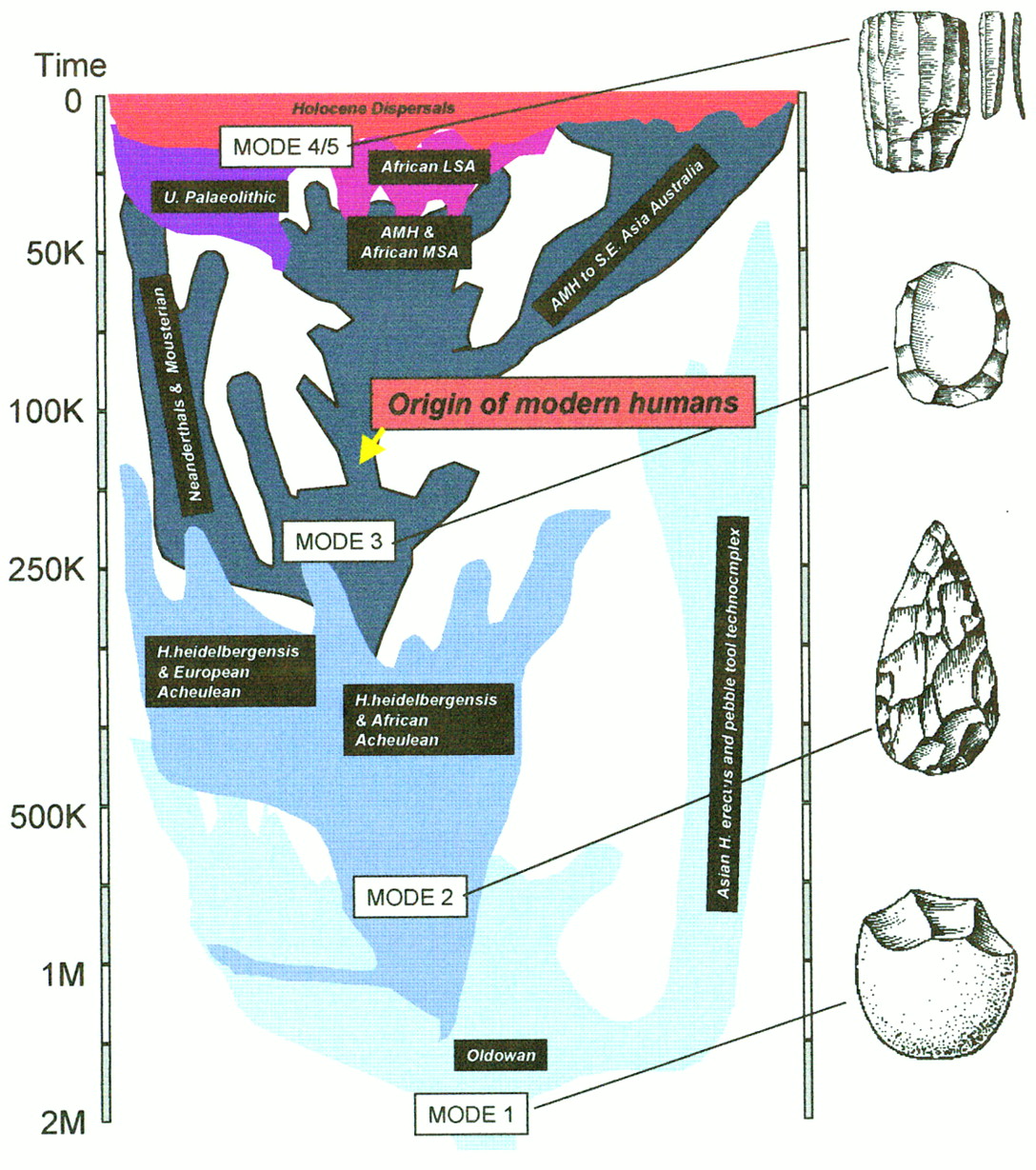
Finally, there has been a tendency to collapse a number of separate problems together: coalescence times of genes, divergence and dispersal of populations, and the evolution of anatomical and behavioral modernity. Increasingly, the archeological and fossil records are showing that these may be separate events, and yet to many they are all the “origins of modern humans.” The complexity of both genetic and fossil evidence should remind us that there may be many such “origins” and that they could be scattered temporally and geographically. A move away from simple phylogenetic questions and approaches to human evolution might help us to untangle these various events and thus develop a more integrated evolutionary model of biological and behavioral history. Such a move will require knowing far more about the context of human genetic evolution, and that is what the paleobiological record can best provide.
Notes
[1] E-MAIL ; FAX 44 (0) 1223-335460.
REFERENCES
- ↵P. Brown(1987) Pleistocene homogeneity and Holocene size reduction: The Australian human skeletal evidence. Archaeol. Oceania 22:41–67.
- ↵R.L. CannM. StonekingA.C. Wilson(1987) Mitochondrial DNA and human evolution. Nature 325:31–35.
- ↵L.L. Cavalli-SforzaA. PiazzaP. Menozzi(1994) History and geography of genes. (Princeton University Press, Princeton, NJ).
- ↵R.A. FoleyM.M. Lahr(1992) Beyond out of Africa: Reassessing the origins of Homo sapiens.. J. Hum. Evol. 22:523–529.
- ↵(1997) Mode 3 technologies and the evolution of modern humans. Cambridge Archeol. J. 7:3–36, ibid.
- ↵M. FullertonR.M. HardingR.C. GriffithsJ.B. Clegg(1997) The genetic ancestry of modern humans: Inferences from the analysis of DNA sequence diversity at the beta-globin locus. Am. J. Hum. Biol. 9:118.
- ↵C.P. GrovesM.M. Lahr(1994) A bush not a ladder: Speciation and replacement in human evolution. Perspectives Hum. Biol. 4:1–11.
- ↵R. GrunC.B. Stringer(1991) Electron spin resonance dating and the evolution of modern humans. Archeometry 33:153–199.
- ↵R. GrunP.B. BeaumontC.B. Stringer(1990) ESR dating evidence for early modern humans at Border Cave in South Africa. Nature 344:537–539.
- ↵H. Harpending(1994) Signature of ancient population growth in a low resolution mitochodrial DNA mismatch distribution. Hum. Biol. 66:591–600.
- ↵H. HarpendingS.T. SherryA.R. RogersM. Stoneking(1993) The genetic structure of ancient human populations. Curr. Anthropol. 34:483–496.
- ↵W.W. Howells(1989) Skull shapes and the map. Papers of the Peabody Museum, archeology and ethnology. (Harvard University Press, Cambridge, MA).
- ↵J.J. HublinF. SpoorM. BraunF. ZonneveldS. Condemi(1996) A late Neanderthal associated with Upper paleolithic artefacts. Nature 381:224–226.
- ↵J. KammingaR.V.S. Wright(1988) The Upper Cave at Zhoukoudian and the origins of the Mongoloids. J. Hum. Evol. 17:739–767.
- ↵R.G. Klein(1992) The archeology of modern human origins. Evol. Anthropol. 1:5–15.
- ↵M. KringsA. StoneR.W. SchmitzH. KrainitzkiM. StonekingS. Paabo(1997) Neanderthal DNA sequences and the origin of modern humans. Cell 90:19–30.
- ↵M.M. Lahr(1994) The multiregional model of modern human origins: A reassessment of its morphological basis. J. Hum. Evol. 26:23–56.
- ↵(1996) The evolution of human diversity. (Cambridge University Press, Cambridge, UK) ibid.
- ↵M.M. LahrR.A. Foley(1994) Multiple dispersals and modern human origins. Evol. Anthropol. 3:48–60.
- ↵S.L. LarnachN.W.G. Macintosh(1974) A comparative study of Solo and Australian aboriginal crania. in Grafton Elliot Smith: The man and his work, eds A.P. ElkinN.W.G. Macintosh(Sydney University Press, Sydney, Australia), pp 95–102.
- ↵D. Leiberman(1995) Testing hypotheses about recent human evolution from skulls. Curr. Anthropol. 36:159–197.
- ↵D.R. Maddison(1991) African origin of human mitochondrial DNA reexamined. System. Zool. 40:355–363.
- ↵D.R. MaddisonM. RuvoloD.L. Swofford(1992) Geographic origins of human mitochondrial DNA: Phylogenetic evidence from control region sequences. System. Biol. 41:111–124.
- ↵N. MercierH. ValladasJ.L. JoronJ.L. ReyssF. LevequeB. Vandermeersch(1991) Thermoluminescence dating of the late Neanderthal remains from Saint-Cesaire. Nature 351:737–739.
- ↵M. NeiN. Takahata(1993) Effective population size, genetic diversity and coalescence time in sub-divided populations.. J. Mol. Evol. 37:240–244.
- ↵J. RelethfordH.C. Harpending(1994) Craniometric variation, genetic theory, and modern human origins. Am. J. Phys. Anthropol. 95:249–270.
- ↵W.J. RinkH.P. SchwarczF.H. SmithJ. Radovcic(1995) Ages for Krapina hominids. Nature 378:24.
- ↵A.C. Rogers(1995) Genetic evidence for a Pleistocene population explosion. Evolution 49:608–615.
- ↵A.R. RogersH. Harpending(1992) Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 9:552–569.
- ↵M.E. RuvoloS. ZehrM. Von Dornum(1993) Mitochondrial COII sequences and modern human origins. Mol. Biol. Evol. 10:1115–1135.
- ↵N.J. Shackleton(1996) New data on the evolution of Pliocene climatic variability. in Paleoclimate and neogene evolution, ed E. Vrba(Yale University Press, New Haven, CT), pp 282–290.
- ↵M. StonekingS.T. SherryA.J. ReddL. Vigilant(1992) New approaches to dating suggest a recent age for the human mtDNA ancestor. Phil. Trans. R. Soc. Lond. B 337:167–175.
- ↵C.B. Stringer(1992a) Reconstructing recent human evolution. Phil. Trans. R. Soc. Lond. B 337:217–224.
- ↵G. BrauerF.H. Smith, eds(1992b) Replacement, continuity and the origin of Homo sapiens. Continuity or replacement: Controversies in the evolution of Homo sapiens (Balkema, Rotterdam, Netherlands), pp 9–24, ibid.
- ↵E.S. VrbaG.H. DentonT.C. PartridgeL.H. Burckle, eds(1995) The evolution and distribution of later Pleistocene human populations. Paleoclimate and evolution with emphasis on human origins (Yale University Press, New Haven, CT), pp 524–531, ibid.
- ↵———. 1998. A metrical study of the WLH-50 calvaria. J. Hum. Evol. (in press)..
- ↵C.B. StringerJ.J. HublinB. Vandermeersch(1984) The origin of anatomically modern humans in Western Europe. in The origin of modern humans: A world survey of the fossil evidence, eds F.H. SmithF. Spencer(Alan R. Liss, New York, NY), pp 51–135.
- ↵C.B. StringerR. GrunH.P. SchwarczP. Goldberg(1989) ESR dates for the hominids from Israel. Mitteilungen der Antropologischen Gessellschaft in Wien 7:49–62.
- ↵C.C. SwisherW.J. RinkS.C. AntonH.P. SchwarczG.H. CurtisA. SuprijoWidiasmoro(1996) Latest Homo erectus of Java—potential contemporaneity with Homo sapiens in Southeast Asia. Science 274:1870–1874.
- ↵A.R. Templeton(1992) Human origins and analysis of mitochondrial DNA sequences. Science 255:737.
- ↵(1993) The “eve” hypothesis: A genetic critique and reanalysis. Am. Anthropol. 95:51–72, ibid.
- ↵E. Trinkaus(1990) Cladistics and the hominid fossil record. Am. J. Physical Anthropol. 83:1–11.
- ↵D. TurbonA. PerezC.B. Stringer(1997) A multivariate analysis of Pleistocene hominids: Testing hypotheses of European origins. J. Hum. Evol. 32:449–468.
- ↵G.N. Van Vark(1990) A study of European Upper Paleolithic crania. Fysisch-Antropologische Mededelingen 2:7–15.
- ↵G.N. Van VarkW. Henke(1992) Affinities of European Upper paleolithic Homo sapiens and later human evolution. J. Hum. Evol. 23:401–417.
- ↵L. VigilantM. StonekingH. HarpendingK. HawkesA. Wilson(1991) African populations and the evolution of human mitochondrial DNA. Science 253:1503–1507.
- ↵D.M. Waddle(1994) Matrix correlation tests support a single origin for modern humans. Nature 368:452–545.
- ↵E. WatsonP. ForsterM. RichardsH.-J. Bandelt(1997) Mitochondrial footprints of human expansions in Africa. Am. J. Hum. Genet. 61:691–704.
- ↵Weidenreich, F. 1943. The skull of Sinanthropus pekinensis: A comparative study of a primitive hominid skull. Paleontol. Sinica 10. .
- ↵M. WolpoffR. Caspari(1997) Race and human evolution. (Simon & Schuster, New York, NY).
- ↵M.H. WolpoffX.Z. WuA.G. Thorne(1984) Modern Homo sapiens origins: A general theory of hominid evolution involving the fossil evidence from East Asia. in The origin of modern humans: A world survey of the fossil evidence, eds F.H. SmithF. Spencer(Alan R. Liss, New York, NY), pp 411–483.