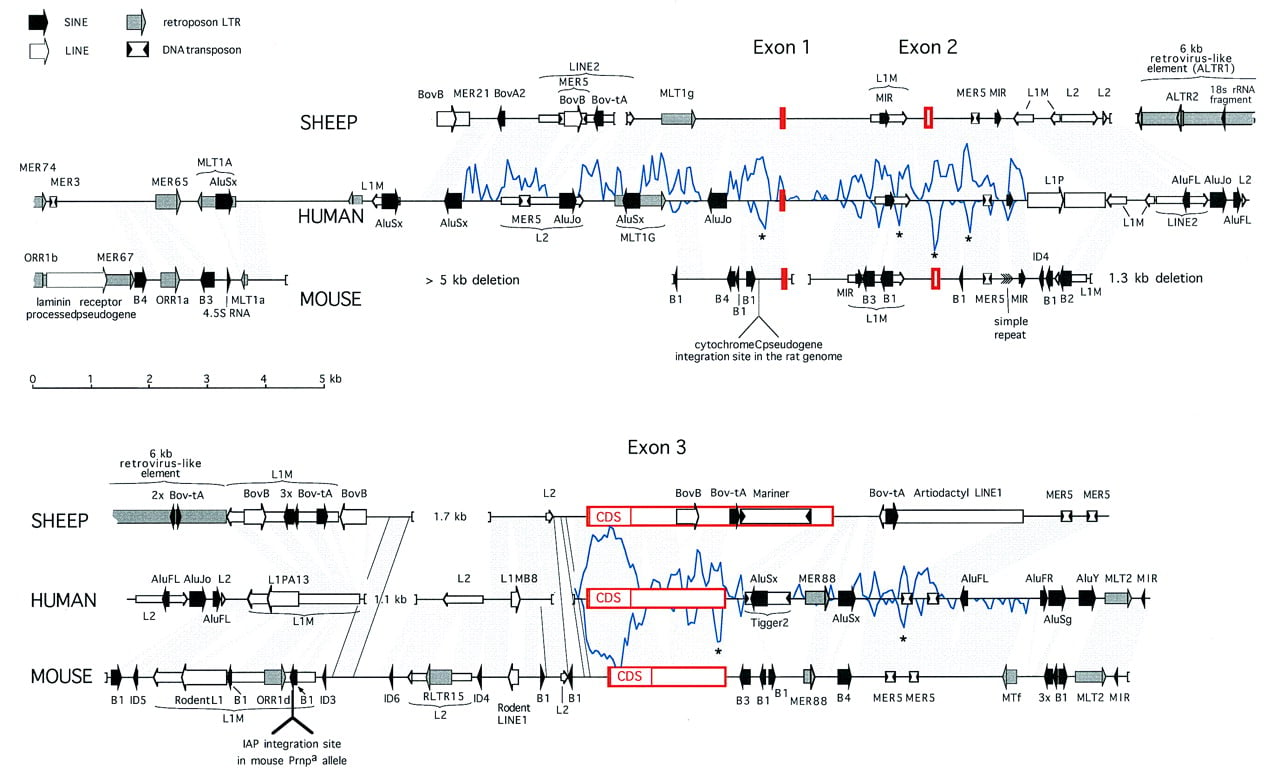
Comparison of the human, sheep, and mouse PrP locus sequences. The complete sheep and human sequences are shown, whereas thePrnpa mouse cosmid sequence extends 2.5 kb upstream of the aligned regions. Sequences orthologous between human and either mouse or sheep are indicated by shaded areas. In two regions deleted in the human sequence, orthologous sequences between sheep and mouse are indicated with outlined shading. Deletions in the genome >250 bp are indicated by brackets. Repeats were identified as described in Methods. Identified interspersed repeats are represented by four differently shaded arrows for the four major classes of repeats: SINEs, LINEs, LTR elements, and DNA transposons. Wide arrows are used for elements that are absent at the orthologous sites in the other two species and thus probably integrated after the mammalian radiation. Older elements are indicated with thinner arrows. The indicated shared interspersed repeats are usually not detectable in the mouse genome (and sometimes not in the sheep genome) by direct comparison with the repeat consensus sequence but could be inferred from the alignment with human DNA. The mariner and large retroviral insert in sheep and the multiple SINEs in mouse are discussed in the text and Figs. 3 and 4. Briefly, other nonfamiliar, but common, inserts are as follows: AluFL andAluFR in human are the older, free left and right arm monomeric precursors of the Alu dimer, Bov-B in sheep is a LINE-like element specific to true ruminants (Szemraj et al. 1995; Smit 1996), and the mammalian-wide LINE2 (L2) elements were responsible for the distribution of the MIR SINEs (A. Smit, unpubl.). All newly derived repeat consensus sequences and descriptions have been incorporated in RepBase Update (Jurka and Smit 1997). The location of the cytochromec pseudogene in rat is based on sequence data of Saeki et al. (1996). A measure of similarity between the human and either sheep or mouse sequences, depicted by a graph above and below the human sequence, respectively, is derived from raw cross_match (Smith–Waterman) scores of 100-bp fragments (overlapping by 50 bp) of human sequence with their orthologous sites in sheep and mouse using a matrix. The lowest score above which virtually all matches had been found to be significant during construction of the RepeatMasker program (A. Smit and P. Green, unpubl.), was used as a cutoff score and subtracted from the total score to form the baseline. Regions outside the coding region that are conserved between all species are indicated with an asterisk (*) and were studied in detail. Alignments of the most interesting regions are shown in Fig. 6.