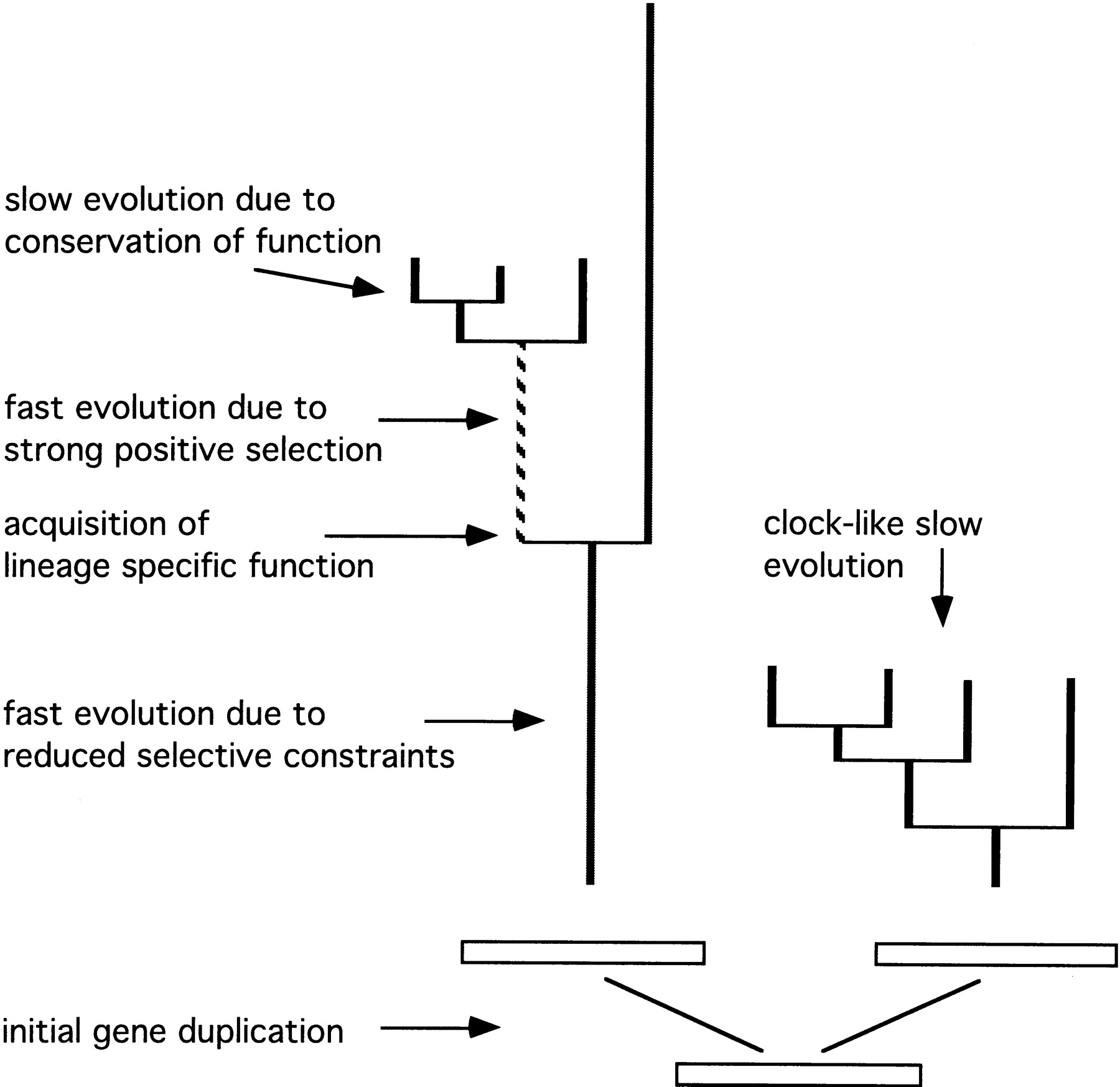
Model for the evolution of orphan genes. The model assumes an initial gene duplication, after which selective constraints in one of the duplicated genes become relaxed. This allows a fast evolutionary divergence (left), indicated by a long branch in the topology. After a lineage splitting event, the gene may become integrated into a new central function in one lineage, but not in the other, where it continues to evolve quickly because of reduced constraints. The new function in the first lineage implies that the gene would go through a phase of adaptive evolution, which would also result in a long branch, depending on how many amino acid changes occurred during the phase of adaptation. But once an adaptive peak is reached, further evolution is slowed down and the branches become short. At this time, the gene may have lost all sequence similarity to its parent gene, but not necessarily its structural similarity. The parent gene (right topology) would undergo the same lineage splitting events, but would continue to have short branches in all lineages, because it has retained its original function. This model suggests the existence of three types of divergence modes: (1) fast divergence of genes which may or may not yet have lost their sequence similarity to their parent gene, (2) fast divergence due to positive selection, and (3) slow-evolving orphan genes. Note that the model would apply in a similar way if the initial gene would not have been created through a pure gene duplication, but through recruitment and recombination of exons from other genes, or even after a gene has lost its original function in the context of a speciation event.